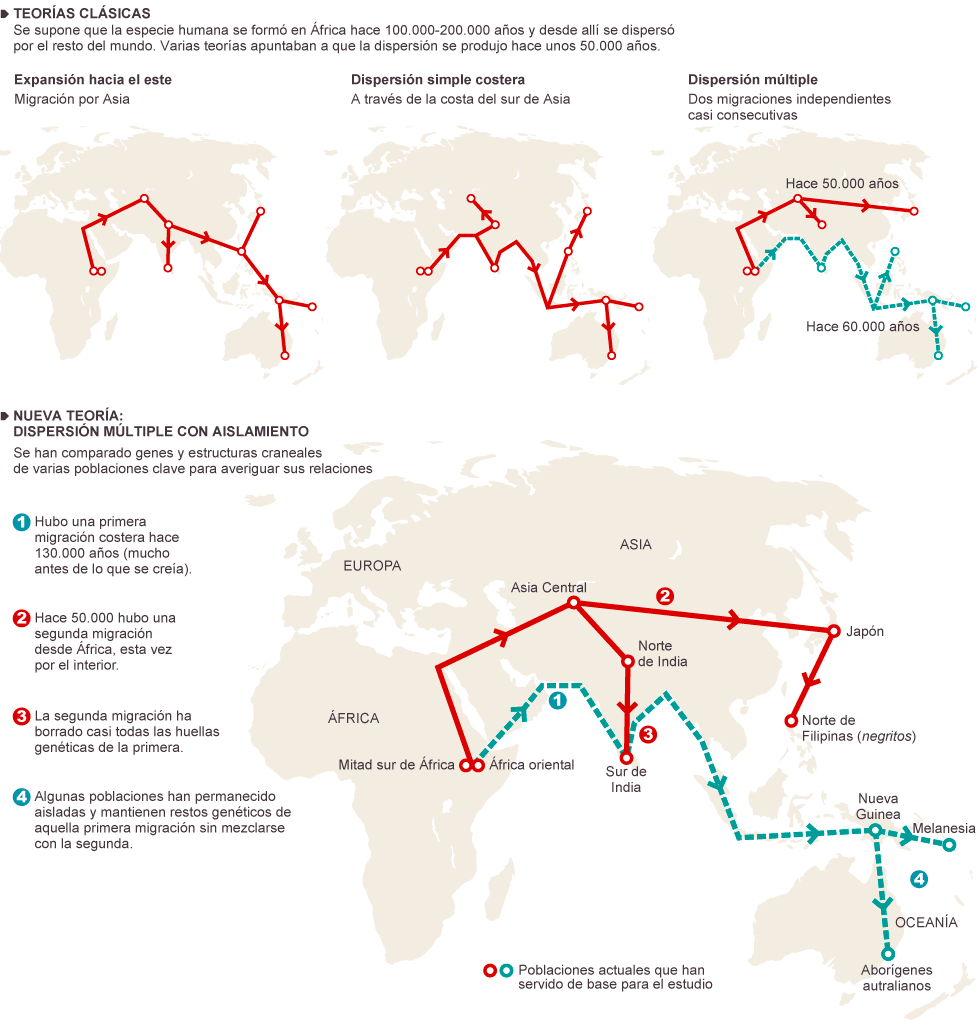
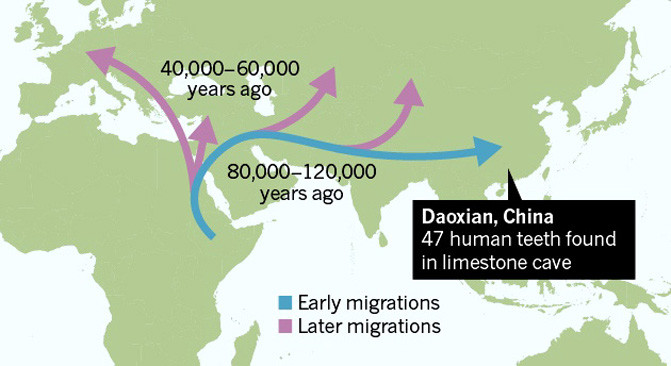
![stone tools infographic 5-20-15]() |
| Primeras tecnologías líticas |
Lomekwiense
La fabricación de herramientas está bien documentada en chimpancés, gorilas, orangutanes y ciertos monos. Con la notable excepción de los martillos de piedra usados por los chimpancés para partir nueces, las herramientas de piedras han sido fabricadas por los Hominini.
La primera producción de herramientas de piedra marca el comienzo de un comportamiento innovador que transformó la ecología, los sistemas sociales y la cultura de los Hominini mediante el aprendizaje social de conocimientos y tecnología (Erella Hovers, 2015).
Los conjuntos de Gona (ca 2,6 Ma), eran más perfectos de lo que podía esperarse para una primera experiencia de fabricación lítica. El descubrimiento de los artefactos de Lomekwi 3 validaron la hipótesis de un origen antiguo.
Pulsa para saber más sobre los útiles prehistóricos, sus procesos de fabricación, su evolución y su uso![]() |
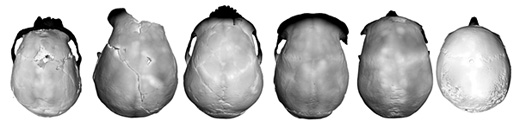
| Localización de Lomekwi 3 (LOM3) y otros sitios arqueológicos al Oeste del Lago Turkana. Harmand et al, 2015. |
Sonia Harmand et al (2015), han informado del hallazgo en 2011-2012 en Lomekwi 3 (LOM3), en el lado occidental del lago Turkana, Kenia, de 149 herramientas líticas (120 en superficie, 10 en la ladera y 19
in situ)
, datadas en 3,3 Ma, 700 ka más antiguas que las conocidas hasta el momento (pero
Domínguez-Rodrigo y Alcalá, 2016).
- 83 núcleos.
- 35 lascas.
- 7 elementos pasivos, potenciales yunques.
- 7 percutores.
- 3 guijarros trabajados.
- 2 guijarros divididos.
- 12 artefactos indeterminados.
Este hallazgo, junto con el de huesos con marcas de corte datados en hace 3,39 Ma en DIK-55, Dikika (
McPherron et al 2010 y
Thompson et al 2015, pero
Manuel Domínguez-Rodrigo, Travis Rayne Pickering y Henry T. Bunn 2011; Domínguez-Rodrigo y Alcalá, 2016), implica que los fabricantes de las primeras herramientas de piedra posiblemente no fueron
Homo y en este caso, el agarre de precisión no fue exclusivo de este género.
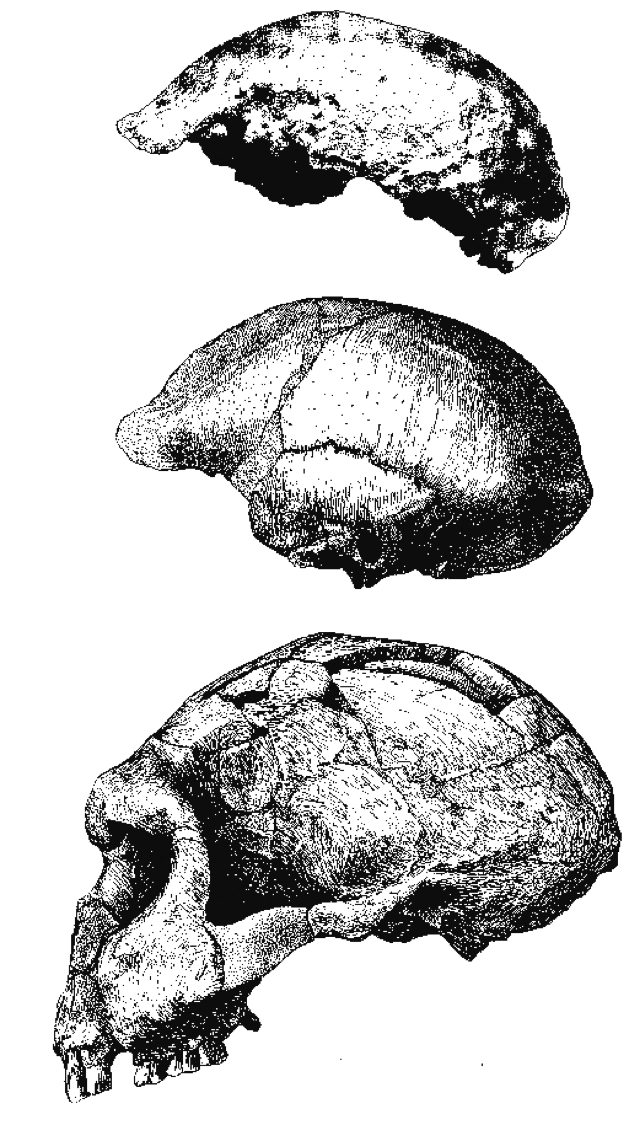
- Estos hallazgos coinciden en geografía y cronología con fósiles atribuidos a Kenyanthropus platyops, mientras que Australopithecus afarensis aparece en el Lower Awash Valley hace 3,39 Ma, en asociación con los huesos con marcas de corte de Dikika. La mandíbula parcial LD 350-1 de Ledi-Geraru es la evidencia más temprana del género Homo, datada en hace 2,8 Ma.
- Yonatan Sahle, Sireen El Zaatari y Tim D. White (2017) advierten de la posibilidad de que las marcas de corte atribuidas a los homininos arcaicos puedieran ser causadas por cocodrilos.
Estas herramientas no se corresponden con la tecnología olduvaiense, dominada por las lascas:
- Alrededor del 76% de los hallazgos en LOM3 son núcleos, yunques, martillos y guijarros trabajados, y los principales modos de talla son el percutor pasivo y el bipolar, lo que indica que las actividades principales se asocian con golpes contra yunques, con movimientos similares a lo que realizan los chimpancés para partir nueces, en lugar de tomar el núcleo en la mano, como en el olduvaiense. Se utilizan dos técnicas:
- El bloque a tallar se mantiene sobre el yunque con una mano mientras la otra utiliza un percutor para obtener las lascas a partir del bloque.
- Se golpea el bloque a tallar directamente sobre el yunque.
- Algunos objetos fueron utilizados como yunques o martillos y como núcleos para producir escamas, lo que apunta a variabilidad en la explotación de los recursos.
- Predominan los núcleos realizados a partir de guijarros de bloques de lava grandes y pesados de origen local. La mayoría de los núcleos fueron obtenidos a partir de una plataforma de golpeo sobre una única superficie, lo que dio como resultado varias extracciones unidireccionales superpuestas y contiguas (explotación parcial unifacial). Unos pocos especímens muestran descamación unifacial multidireccional, mientras que otros muestran descamación bifacial.
- Las muestras de LOM3 son más grandes y pesadas que las posteriores olduvaienses.
- Las escamas son más masivas que las producidas accidentalmente por los chimpancés, pero los yunques y martillos están dentro del rango de tamaño de los utilizados por los chimpancés. A menudo, las lascas se obtuvieron de forma exitosa, pero también son visibles accidentes significativos de talla. La precisión de las percusiones fue ocasionalmente pobremente controlada, como muestran las marcas repetidas causadas por golpes fallidos.
Por ello los autores sugieren la nueva etiqueta Lomekwiense para referirse a esta primera tecnología lítica, que representa una fase intermedia entre un hipotético uso de herramientas de piedra para golpear con ellas y la talla para obtener lascas.
![]() |
Artefactos de LOM 3.
a. Elemento pasivo, yunque. In situ. 12 kg.
b. Elemento pasivo, yunque. 4,9 kg.
Ambos exhiben similares marcas cruzadas de fracturas, aplastamientos e impactos. En a, el daño está localizado en una cara lateral, con las marcas en un plano horizontal. En b, el daño está distribuido a lo largo de la mayor parte del perímetro, sin marcas percusivas en el plano horizontal. En ambos casos, la intensidad de las marcas indica un uso en trabajos pesados.
c. Núcleo unifacial. 4,74 kg. Técnica bipolar y explotación semiperiférica. El recuadro muestra marcas de aplastamiento en la superficie proximal.
Harmand et al, 2015 |
![]() |
Artefactos de LOM3.
a. Elemento pasivo, yunque. Pesado bloque rectangular con una morfología natural que le proporciona estabilidad.
b. Martillo de piedra que muestra impactos aislados. 3,09 kg.
c. Martillo de piedra que muestra impactos aislados y una fractura en la terminación.
Harmand et al, 2015 |
![]() |
Artefactos de LOM3. a. Núcleo in situ de 1,85 kg y lasca retocada de 650 g. Núcleo unifacial, percutor pasivo y técnica bipolar. Tanto el núcleo como la lasca muestran marcas de percusión dispersas que indican que las actividades de percusión se realizaron antes de remover la lasca y que el bloque fue utilizado para diferentes propósitos. b. Núcleo unifacial in situ, técnica bipolar. 3,45 kg. c. Núcleo unifacial. Técnica percutor pasivo. 1,84 kg. |
![]() |
Dibujo sobre material procedente de Gona EG 10 y EG 12. 1 a 7, núcleos con diferentes tipos de explotación. 8 a 10, lascas. Semaw (2000). |
Preolduvaiense
La garganta de Olduvai ha dado nombre a la industria lítica olduvaiense (complejo industrial olduvaiense) caracterizada por los pocos golpes necesarios para obtener los útiles: uno solo en ocasiones. El resultado es el de unas herramientas más bien burdas, pero con una tosquedad que engaña. Ni resulta fácil saber dar golpes precisos para obtener los filos cortantes, ni se puede despreciar el filo de las lascas que se desprenden del núcleo cuando se domina la técnica de una forma aceptable. Los artefactos se manufacturaron a partir de rocas que provenían de diferentes lugares a algunos kilómetros de distancia.
Principales caracteres distintitivos:
- Principalmente herramientas de corte que pertenecen a procesos de descamación.
- La frecuencia de martillos y otras herramientas de percusión es extremadamente baja (por ejemplo, Kibunjia 1994; Roche et al, 1999; Semaw 2000; Delagnes y Roche 2005). Los elementos de percusión faltan en Gona, Hadar y LA1 en Turkana Occidental. Cuando se encuentran, como en LA2C, documentan movimientos de percusión directa altamente controlados. Los martillos muestran una alta densidad de cicatrices de impacto en áreas circunscritas, que se produjeron por un uso preciso y recurrente, según hábitos motores estables (Delagnes y Roche 2005).
- Conocimiento avanzado de la mecánica de la fractura de piedra
- Percusión directa, rara vez percusión bipolar.
- Está escasamente documentada la percusión dedicada a actividades distintas de la talla.
El surgimiento de la manufactura lítica pudo estar asociado con la aridez y el aumento de espacios abiertos, aunque luego se extendiera su uso a todo tipo de hábitats. Las ventajas de esta innovación tuvieron que ser mayores que el coste de obtención y transporte. Debió de ser esencial el acceso a los recursos nutritivos en el interior de los huesos o en el encéfalo, con revestimiento óseo y la ampliación del repertorio de plantas consumibles. Las condiciones debieron favorecer estrategias sociales consistentes en el transporte de carcasas y herramientas a determinados puntos del paisaje, ya sea un campamento central o múltiples lugares de forrajeo (Potts, 2013).
Los primeros sistemas técnicos humanos, datados en más de 2 Ma, muy simples se hallaron en los siguientes yacimientos:
![]() |
| Yacimientos etíopes. |
Formación Hadar, Middle Awash (Etiopía).
Los primeros hallazgos de Gona se produjeron al principo de los 1970 (Johanson et al. 1978, 1982). Al este del río Kada Gona aparecieron dispersiones de baja densidad de artefactos superficiales (Corvinus 1976; Corvinus y Roche 1976, 1980; Roche y Tiercelin 1977, 1980). Las excavaciones posteriores en West Gona permitieron descubrir artefactos in situ de baja densidad (Harris 1983; Harris y Semaw 1989). Las excavaciones sistemáticas y los análisis geocronológicos comenzaron en 1992–1994. Más localidades a lo largo de los ríos Kada y Ounda Gona produjeron artefactos de superficie e in situ, en su mayoría recuperados de EG10, EG12, y OGS-7. EG10 y EG12 produjeron altas densidades de artefactos de piedra sin huesos asociados (Semaw 2006). OGS-7 es el único sitio, y el más antiguo, donde se descubrieron artefactos con huesos fósiles, algunos de los cuales posiblemente muestran modificaciones humanas (Semaw et al. 2003). Huesos modificados de una edad similar que muestran marcas de corte y percusión hechas por herramientas de piedra fueron descubiertos en Bouri en el Medio Awash, pero sin ningún artefacto asociado (de Heinzelin et al. 1999). Los tres conjuntos líticos están mínimamente perturbados. Se centran exclusivamente en la producción de escamas pequeñas y medianas a partir de un alto grado de selectividad de la materia prima, basada en el tipo de roca, los fenocristales y la masa de fondo (Stout et al. 2005).
La técnica de tallado es percusión directa con martillo duro, sin evidencia de técnicas bipolares, de yunque o de lanzamiento. Se observaron cinco métodos de descamación: unifacial simple, unifacial centrípeto, unidireccional, bifacial parcial y multifacial irregular. Sin embargo, el conjunto EG10-12 está dominado por núcleos unifaciales. En OGS-7, los núcleos son bifaciales o multifaciales, con un mayor número de cicatrices en escamas que indican una reducción relativamente intensa. Según Stout et al. (2010), aunque no se puede descartar la existencia de distintas tradiciones tecnoculturales, esta variabilidad ha sido interpretada como posiblemente originada por la diversidad ambiental. OGS-7 se ubica en un banco de canal o margen de canal, mientras que tanto EG-10 como EG-12 se localizan en una llanura de inundación (Stout et al. 2010).
En Hadar, A.L. 666 produjo un conjunto lítico muy restringido de escamas y fragmentos angulares, y representa la co-ocurrencia espaciotemporal más antigua de herramientas, fauna y Homo.
En A.L. 894, se descubrieron unos pocos cientos de fragmentos de huesos y varios miles de artefactos líticos en los depósitos de arcilla limosa de la llanura de inundación de una corriente de baja energía. La fauna de roedores proporciona evidencia importante de un cambio de fauna entre 3,2 Ma y 2,4 Ma. Este cambio documenta la aridez creciente, pero no un cambio dramático en el paleoambiente local. Este cambio coincide también con la extinción local de Australopithecus afarensis y la aparición del género Homo en Hadar.
El conjunto lítico representa posiblemente el palimpsesto de varias ocupaciones. La interpretación tafonómica del conjunto de huesos bien conservados muestra la falta de asociación funcional entre los artefactos y los huesos, lo que subraya la complejidad de los procesos de formación del sitio (Domínguez-Rodrigo y Martínez-Navarro 2012). Sin embargo, las numerosas modificaciones entre los elementos líticos sugieren que el entierro fue relativamente rápido y con una perturbación geológica mínima (Hovers 2009). La mayoría de los artefactos son escamas completas o rotas (83%), con núcleos en números pequeños (1%). Los fragmentos angulares forman la parte restante del ensamblaje. En lugar de vincularse a escamas antrópicas, bien podrían haber sido el resultado de mecanismos de post-deposición (Hovers 2003).
Se utilizaron como materias primas rocas volcánicas, principalmente riolita, basalto y traquita. El tamaño de los guijarros sugiere transporte desde una fuente ubicada al menos a varios metros de distancia. Fueron seleccionados con precisión en el conglomerado cercano, especialmente en el caso de las riolitas. Sin embargo, en A.L. 666, las rocas son más homogéneas y de grano más fino que en A.L. 894. También se explotaron el cuarzo y el chert. No se puede descartar el transporte desde una fuente desconocida. Estos patrones sugieren no solo la selectividad, sino también posiblemente la búsqueda de materias primas de la mejor calidad. Además, algunos guijarros se importaron al sitio parcialmente decorticados, después de que los perceptores los probaran en la fuente (Goldman-Neuman y Hovers 2009, 2012). La descamación en A.L. 894 fue principalmente unipolar, mientras que en A.L. 666 se documentó la descamación bipolar para la explotación de cuarzo y quert. Las cicatrices revelan errores de talla e intentos de rectificado mediante giro de los núcleos (Hovers 2009; Goldman-Neuman y Hovers 2012).
YACIMIENTO | DATACIÓN | OBJETOS | MATERIAL | ASOCIACIONES | OBSERVACIONES |
Kada Hadar. Kada Gona 2.3.4 | 2,6 Ma | 15 | traquita y basalto |
| Recuperados en la segunda mitad de los setenta. |
Kada Hadar. Kada Gona EG10 | 2,6-2,52 Ma | 2.236 en excavación de 13 m2 | traquita y riolita, local |
|
|
Kada Hadar. Kada Gona EG12 | 2,6-2,52 Ma | 309 en superficie y 444 en excavación de 8 m2 | traquitay riolita, local |
|
|
Ounda Gona OGS6 y OGS7 | 2,6 Ma | 200 en superficie y 500 en excavación de 2,6 m2 | materiales volcánicos vítreos y afanitas de gran calidad, local y no local | Fauna. | Conjunto con trazas de despiece más antiguo conocido |
Localidad AL666 | 2,33 Ma | 224 | Basalto y chert. Local y no local | Homo habilis y herbívoros | Trazas de despedazamiento |
YACIMIENTO | CADENA OPERATIVA |
Kada Hadar. Kada Gona 2.3.4 | 4 cantos unifaciales. Dos de ellos presentan las extracciones en el lateral del eje mayor. 2 cantos bifaciales. 3 poliedros parciales con extracciones en la dirección de la anchura. 2 núcleos. |
Kada Hadar. Kada Gona EG10 | En su mayor parte, bases positivas unifaciales y fragmentos. Nucleos, lascas, escombro, martillos. Sin retocar. |
Kada Hadar. Kada Gona EG12 | En su mayor parte, bases positivas unifaciales y fragmentos. Nucleos, lascas, escombro, martillos. Sin retocar. |
Ounda Gona OGS6 y OGS7 | Peso importante de restos de talla. Lascas, núcleos muy agotados, escombros. |
Localidad AL666 | La mayoría son bases positivas. 3 choppers transversales bifaciales recogidos en superficie. 1 núcleo recogido en excavación, con levantamientos desde diferentes plataformas de percusión. |
Además:- Kada Gona EG 13: Un hueso con marcas de corte.
- Kada Gona WG 9: Dos huesos con marcas de corte.
- Localidad AL849: 2,4-2,3 Ma. 20 m2. Homo habilis. 4.828 objetos de material local, a partir de método unifacial a mano alzada.
Gona: Kimbel et al (1996). Semaw et al (1997, 2002, 2003). Domínguez-Rodrigo et al (2005). Stout et al (2005, 2010). Semaw, 2006.
Hadar: Hovers (2003, 2009) Goldman-Neuman y Hovers (2009, 2012).
Formación Omo-Shungura (Etiopía).
De acuerdo con la descripción tipológica de Chavaillon (1976), la colección de Omo está compuesta de ragmentos, escamas y núcleos de cuarzo de pequeño tamaño y de desechos abundantes. Estos últimos se consideraron como residuos naturales, desprendidos durante la descamación, o como fragmentos de piedra de martillo producidos por percusión intensa. Debido a la falta de tipos líticos definidos en Olduvai, Chavaillon (1976) introdujo el término “Shungura facies” para las industrias de Omo.
Howell et al. (1987) discutieron el contexto y la posición estratigráfica de Omo 71 y Omo 84, los dos sitios pertenecientes al Miembro E. Más tarde, de la Torre (2004) cuestionó la evidencia de intencionalidad en los artefactos interpretándolos como ensamblajes naturales. Esto se confirmó durante una investigación reciente en la Formación Shungura (Delagnes et al. 2011).
En el miembro F, se descubrieron cinco sitios arqueológicos, en particular FtJi 1, 2, 5, Omo 57 y Omo 123. Merrick y Merrick (1976) los dividieron en sitios en posición primaria (FtJi 2 y Omo 123), y sitios en contexto secundario del canal fluvial (FtJi 1, 5 y Omo 57). De acuerdo con la descripción disponible, las industrias de cuarzo de FtJi 1, 2 y 5 incluyen pocas escamas enteras y rotas y un alto número de fragmentos angulares (Merrick y Merrick 1976). Según Chavaillon (1976), además de escamas y fragmentos angulares, los ensamblajes Omo 57 y 123 también contenían núcleos. Esto fue confirmado por de la Torre (2004), quien señaló, sin embargo, que muchos de los llamados artefactos en realidad son objetos no trabajados. Reconoció la explotación unidireccional unifacial sistemática de los núcleos y, en ocasiones, incluso una rotación destinada a aprovechar al máximo las superficies de talla. Dado que las escamas muestran extirpaciones previas de la cara dorsal, se desprendió de los núcleos más de una serie de escamas. Sin embargo, el pequeño tamaño de los guijarros de cuarzo angular utilizados como positivos no permitió una alta productividad de la escama. Los ángulos naturales y las superficies disponibles se utilizan hasta el agotamiento, sin ningún intento de rejuvenecimiento. A pesar del pequeño tamaño de los positivos, fue ampliamente utilizado el martillo directamente y la técnica bipolar se adoptó solo en dos casos. Se registran muy pocos accidentes de talla, lo que sugiere cierto control de la fuerza utilizada para impactar los núcleos (de la Torre 2004).
Las nuevas investigaciones en la Formación Shungura (Boisserie et al. 2008, 2010; Delagnes et al. 2011) permitieron descubrir varias localidades nuevas en el Miembro F, desde pequeñas concentraciones con pocas piezas hasta sitios con abundante material. Los artefactos son casi exclusivamente escamas y fragmentos de cuarzo. Los estudios sistemáticos demostraron que los guijarros de cuarzo disponibles en los paleocanales cerca de los sitios arqueológicos no superan en número a ningún otro tipo de roca, como lavas, granitos y rocas criptocristalinas. Viceversa, el cuarzo domina en las colecciones arqueológicas. Esto implica una selección deliberada (Delagnes et al. 2011).
YACIMIENTO | DATACIÓN | OBJETOS | MATERIAL | ASOCIACIONES | OBSERVACIONES |
Omo 57. | 2,36-2,34 Ma | 223. De ellos, 30 en excavación | Cuarzo. Escasa presencia de calcedonia y cuarcita | Restos óseos.
Homo sp. | Guijarros angulares de pequeño tamaño |
Omo 57. FtJi-5 | 2,36-2,34 Ma | 24 in-situ y 77 en superficie | Cuarzo. |
| Arrastre de materiales por el curso fluvial. Intervención antrópica sucesiva con intervalos temporales cortos. |
Omo 84. | 2,36-2,34 Ma | 200 in situ y 90 en superficie | Cuarzo. |
|
|
Omo 71 | 2,36-2,34 Ma | 24 | Cuarzo. |
|
|
Omo 123. | 2,36-2,34 Ma | 1.314 | Cuarzo. Local. | Restos óseos raros.
Paranthropus aethiopicus | Guijarros angulares de pequeño tamaño |
Omo 204. FtJj-1 | 2,36-2,34 Ma | 375 in-situ agrupados en seis pequeños agregados densamente concentrados. Otros 270 en superficie. | Cuarzo. 7 objetos de chert y 2 de lava. | Restos óseos.
|
|
Omo 396. FtJj-2 | 2,36-2,34 Ma | 224 instrumentos in-situ y 131 en superficie. | Cuarzo. 1 objeto de lava. |
| El cuarzo tuvo que ser intoducido por los hominini. Fue propuesto como lugar de ocupación. |
YACIMIENTO | CADENA OPERATIVA |
Omo 57. | Mayoría de pequeños fragmentos. Pocas lascas, núcleos y fragmentos de núcleo.
Método unifacial unidireccional.Mano alzada.
0,8% de retocados. |
Omo 57. FtJi-5 | Mayoría de fragmentos angulares. Pocas lascas estrechas. 3 núcleos hallados en superficie. |
Omo 84. | 72 fragmentos angulares. 66 lascas, la mitad, completas. Fragmentos de núcleos. Muy pocos objetos retocados |
Omo 71. | Fue tallado un canto de pequeñas dimensiones, levantado cinco lascas en las dos caras y convirtiéndolo en un chopper. |
Omo 123. | En su mayoría, fragmentos angulares y bases positivas. 12 núcleos in-situ y 11 en superficie. Método bipolar en dos núcleos.
Método unifacial unidireccional. |
Omo 204. FtJi-1 | En su mayoría, pequeños fragmentos. Además, 23 lascas y fragmentos de lasca y un núcleo. |
Omo 396. FtJi-2 | En su mayoría, pequeños fragmentos. 3 bases positivas enteras y 4 fragmentadas.
|
Howell et al (1987). De la Torre (2004).
Lokalalei, West Turkana, Kenia.
Los niveles arqueológicos produjeron restos de fauna, que están pobremente conservados y carecen de cualquier rastro reconocible de acción humana, excepto de una única marca de corte en un fragmento de hueso recuperado en superficie (Roche et al. 1999; Brugal et al. 2003). Se encontró un diente atribuido a un Homo temprano en Lokalalei 1a, en la misma unidad litoestratigráfica que LA1 (Prat et al. 2005).
Las materias primas están constituidas por lavas (fonolita, traquita, basalto y riolita) disponibles en paleocanales a una distancia máxima de 50 m (Harmand 2009).
En LA1, la descamación se produce principalmente en grandes guijarros, que están redondeados y, por lo tanto, carecen de superficies planas o de plataformas de golpeo naturales adecuadas. Las secuencias de talla resultantes son oportunistas y heterogéneas. Los núcleos muestran evidencias de frecuentes accidentes de rotura y de repetidos daños por impacto de percusiones fallidas (Delagnes y Roche 2005).
LA2C ha sufrido escasa perturbación. Las estrategias técnicas se basan en la implementación de reglas técnicas constantes, que documentan la destreza manual avanzada. Un conjunto de núcleos muestra de una a tres cicatrices de escamas no organizadas (descamación simple), como si se hubieran probado y descartado. Otro conjunto muestra evidencia de descamación organizada en clastos de fonolita de grano fino. Los guijarros completos y los bloques angulares se amontonaron sin ninguna preparación. En dos casos, se realizó una fase preliminar de descamación fuera del sitio. Los núcleos que mostraban descamación organizada estaban principalmente escamados en una sola superficie, que es la más grande disponible. Varias series de escamas fueron extraídas de plataformas naturales o rectificadas. Los numerosos cambios en la dirección de descamación aseguraron que la superficie se mantuviera razonablemente plana y regular. En algunos casos, se realizaron unas pocas eliminaciones en otra cara en una etapa de reducción final. También se documenta la descamación alternativa: para producir varias series de escamas, algunos núcleos muestran dos o tres superficies utilizadas en sucesión como superficies de escamación y como plataformas. Al utilizar y mantener los ángulos disponibles en las superficies, los núcleos resultantes tienen una forma similar a la del primitivo. Algunos núcleos muestran evidencia de retoque en los bordes con un ángulo cercano a 90 ° para su uso posterior como herramienta. En muy pocos casos, el retoque también se produce a lo largo de uno o, más raramente, de dos bordes. Varios guijarros muestran áreas densamente trabajadas por percusión. Son de la misma fonolita de grano medio que los especímenes no trabajados, pero son más pesados. Por lo tanto, es probable una selección de los materiales (Delagnes y Roche 2005).
Para Roche y colaboradores (1999) la técnica utilizada en LA2C implica una preparación y utilización de los materiales muy cuidadosa, con repetición de un esquema claro y estándar y precisión de los gestos, inimaginable con anterioridad para unos hominini tan antiguos y obliga a plantearse que sus capacidades cognitivas debían estar bastante más desarrolladas de lo que se supone por lo común. Un núcleo era golpeado hasta 20 veces para extraer las esquirlas y la elección cuidadosa de los materiales implica que quienes los trabajaban conocían sus propiedades mecánicas. De algunos guijarros se obtuvieron más de 30 lascas, lo que implica un esquema cognitivo complejo y un gran dominio de los gestos.
Para Steele (1999) no podemos ir más allá que atribuirlos a Homo habilis por su mayor capacidad craneal, frente a la del Paranthropus boisei.
YACIMIENTO | DATACIÓN | OBJETOS | MATERIAL | ASOCIACIONES | OBSERVACIONES |
LA1 (GaJh 5) | 2,34 Ma | 445 en 60 m2 | Local | Muy pocas trazas de despiece. Asociación con restos de tortuga y huevos de avestruz.
Homo habilis | Guijarros angulares y globulares de gran tamaño |
LA2C | 2,24 Ma | 2.067 en excavación en 17 m2 y 516 en superficie. | Diverso. Son dominantes las lavas. | Muy pocas trazas de despiece. Asociación con restos de tortuga y huevos de avestruz.
Paranthropus aethiopicus | El yacimiento más rico del Modo 0. Abundantes remontajes. Se documenta la cadena de reducción lítica más antigua de veinte unidades que pueden remontarse. No hay transporte, ni siquiera de materias primas.
Guijarros angulares y globulares de gran tamaño. |
YACIMIENTO | CADENA OPERATIVA |
LA1 | La mayoría de las cadenas son muy cortas. Otras, medianamente amplias. Pocas bases positivas (73) en relación a los demás yacimientos de la misma cronología. Buena colección (50) de bases negativas de explotación, muy arcaicas. 282 fragmentos. Baja frecuencia con respecto al número total de núcleos. Numerosas series de extracciones sin una dirección preferencial ni ordenadas y con numerosos accidentes de talla que parecen el resultado de una manufactura expeditiva y una baja capacidad cognitiva.
Explotación unifacial (principalmente) bifacial, multifacial y multidireccional a mano alzada. |
LA2C | Cadena operativa completa. Elementos de percusión presentes. 90% de productos de talla: Alta frecuencia (49,44%) de objetos de tamaño inferior a 1 cm. Lascas de más de 1 cm 40,63%, una frecuencia mucho más alta que en los demás registros. Morfologías claras y bien conseguidas. 3,33% objetos de formato grande configurados sobre canto. 2,5% de objetos retocados. Mayor frecuencia de núcleos (3,06%) y lascas enteras y menor de fragmentos. 3,09% objetos no transformados.
Explotación simple, unifacial (principal) multidireccional, bifacial y multifacial a mano alzada. |
Kibunjia et al (1992). Kibunjia (1994). Roche et al (1999, 2003). Brown y Gathogo (2002). Delagnes y Roche (2005). Harmand (2009).
Otros yacimientos.
- Dana Aoule (Etiopía). Domínguez-Rodrigo et al, 2005
- DAN-1. 2,6-2,5 Ma. Dos horizontes con artefactos de piedra.
- DAN-2. 2,27-2,1 Ma. Cientos de artefactos en superficie y numerosos en excavación. Marcas de corte en cinco huesos.
- Bouri-Hata, Awash (Etiopía): 2,5 Ma. Cortadores y lascas utilizados para descarnar huesos de antílopes y caballos, probablemente por Australopithecus garhi (deHeinzelin et al, 1999).
- Busidima (Etiopía). 2,5-2 Ma. Núcleos. Esocombros. Domínguez-Rodrigo et al, 2005.
- Senga 5, Río Semliki (R. D. Congo): 2,3?-2? Ma. 435 artefactos obtenidos en excavación y 292 en superficie. Nucleos, escamas y escombros. 97% cuarzo. Obtención local. Numerosos huesos de animal asociados con marcas de corte?. Harris et al, 1987.
- Transvaal. Más de 2 Ma.
- Sterkfontein M5A. 3.246 artefactos. Materia prima local. Asociado a Homo habilis?. Kuman, 2003.
- Wonderwek Cave.
- Malapa.
- Kroomdrai B. 2 artefactos de piedra. Materia prima local. Kuman, 2003.
- Swartkrans, Miembro 1. 17 huesos para excavar. 402 artefactos de piedra. Materia prima local. Asociado a Paranthropus robustus, Homo. Brain, 1993, Clark, 1993. Kuman, 2003.
Características.
- Los yacimientos presentan un patrón episódico, dentro de un intervalo estratigráfico y un rango geológico muy concretos.
- Las rocas se transportan para su escamación intencional y utilización en el procesamiento para la obtención del alimento. Hay un uso selectivo de materias primas.
- Hasta 2,3 Ma, el desplazamiento es de solo unas decenas o centenas de metros.
- Entre 2,3-2 Ma, llega a unos pocos kilómetros.
- Muy simple, arcaico y poco diversificado. Baja frecuencia de objetos retocados. Intervenciones poco sistemáticas.
- Falta de objetos finalistas elaborados sobre canto y en formatos grandes, como los clásicos choppers olduvaienses.
- Conjuntos reducidos, poco densos y con objetos de pequeña dimensión.
- Esquema de talla dominante centrípeto. En AL666, se da rotación del núcleo durante la secuencia de producción. Bien desarrollado sentido de la mecánica de fractura y planificación del proceso de escamación.
Semaw y colaboradores atribuyen el mayor tamaño de los núcleos, con respecto a los posteriores a 2 Ma, a la escasa distancia del yacimiento respecto a los lugares de obtención del material. Rechazan la necesidad de proponer una industria pre-olduvaiense e indican que la cultura de Olduvai tuvo una estasis de al menos 1 Ma.
Bibliografía