HOMO EN EUROPA ENTRE 1,3 Y 0,73 MA | ||
PERIODO MA | FÓSILES | TECNOLOGÍA |
1,4-0,9 | No ha sido identificada la especie, que se denomina, provisionalmente, especie ñ | Modo 1 muy pobre |
0,9-0,73 | Homo antecessor | Modo 1 evolucionado |
0,7-0,615 | Homo antecessor en proceso de extinción. Oleadas migratorias sin éxito demográfico. | Modo 1 Modo 2 pobre |
0,615-0,2 | Grado Heidelbergensis y preneandertales | Modo 2 pleno |
0,2-0,027 | Homo neanderthalensis | Modo 2 Modo 3. Musteriense. |
¿Preneandertal? | ¿heidelbergensis? | |
MIS15-MIS13 621-478 ka | Mauer, Boxgrove | |
MIS12 478-424 ka | Arago | |
MIS11 424-374 ka | Sima de los Huesos | Ceprano, Aroeira |
MIS10-MIS9 374-300 ka | Swanscombe | |
MIS8 300-243 ka | Atapuerca Galería | Bilzingsleben |
MIS7 243-191 ka | Pontnewydd, Steinheim, Ehringsdorf, Tourville, Saccopastore, Petralona |
EUROPA CENTRAL
Heidelberg Man, Mauer Jaw, Descubierto por trabajadores de una cantera en 1907, cerca de Heildelberg (Alemania). Descontextualizado de su posición estratigráfica, su edad es incierta, pero se ha estimado en hace 609 ka (Wagner et al, 2010). Se trata de una mandíbula inferior, ramas ascendentes muy anchas, con el mentón retraído y todos los dientes. Escotadura sigmoideia apenas deprimida, sínfisis alta y retraída. Muelas relativamente pequeñas, sobre todo M3. La mandíbula es extremadamente larga y robusta, como la del Homo erectus, pero con los dientes más pequeños. Es muy difícil interpretar el espécimen desde una perspectiva filogenética.
Schöningen 13/II sedimentary sequence 4 (Spear Horizon) es conocido por sus importantes hallazgos. En 1995 se descubrieron numerosos artefactos de madera y los esqueletos de unos 20-25 caballos descuartizados. No está clara la forma en que los materiales fueron depositados (Mareike C. Stahlschmidt et al, 2015). Se ha estimado una datación máxima de 337-300 ka (Daniel Richter y Matthias Krbetschek, 2015).
 |
| Mandíbula de Mauer. Los dos premolares izquierdos se extraviaron en los 1940. Foto: K. Schacherl. Wagner et al, 2010. |
Bilzingsleben (Valle Wipper, Turingia, Alemania). Fragmentos de al menos tres individuos, asociados a industria de modo 1 evolucionado (micro-Clactoniense) y fauna. Han podido ser reconstruidos dos cráneos. Sin rasgos neandertales. Vlcek (1989) y Mania et al (1994) los asignan a erectus. 0,3-0,28 Ma (Mania y Thomae, 2006). Ambiente de bosque con un clima ligeramente más templado que el actual. Aparecieron más de 10.000 huesos de animales, algunos claramente grabados.También tres estructuras de hábitat y organización del espacio.
· La zona más cercana al agua aglutina los instrumentos líticos, restos de fauna, tres agujeros de poste y defensas de elefante.
· La zona más alejada, aparentemente empedrada, reúne los yunques para machacar huesos, grandes cráneos de bóvidos, maderas y restos de hogares.
Schöningen (Elba-Saale, Alemania) contiene al menos 20 sitios arqueológicos de preservación excepcional en un área de unos 9.400 m2 (Serangeli et al, 2015), con una sucesión sedimentaria de hasta 45 m de espesor (Lang et al, 2015). |
| Algunas piezas obtenidas en Schöningen. Schoch et al, 2015. |
Schöningen 13/II sedimentary sequence 4 (Spear Horizon) es conocido por sus importantes hallazgos. En 1995 se descubrieron numerosos artefactos de madera y los esqueletos de unos 20-25 caballos descuartizados. No está clara la forma en que los materiales fueron depositados (Mareike C. Stahlschmidt et al, 2015). Se ha estimado una datación máxima de 337-300 ka (Daniel Richter y Matthias Krbetschek, 2015).
- Indicios de fuego, pero sin pruebas convincentes de uso humano (Mareike C. Stahlschmidt et al, 2015; Utz Böhner, Jordi Serangeli y Pascale Richter, 2015).
- Nueve lanzas (de empuje), una jabalina (para lanzar), un palo con punta por ambos lados y un palo puntiagudo quemado, obtenidas del tronco de una pícea, dándoles forma con cuidado, tallando las puntas en la madera de la base del tronco, particularmente dura. Se pueden observar en detalle las huellas de corte, raspado y alisado. Están extremadamente bien conservadas y muestran poco o ningún signo de alteración tafonómica. La lanza X fue reafilada tras un uso repetido. El centro de gravedad estaba situado en tercio anterior del arma. Para Robin Dennell estas lanzas indican un grado muy notable de planificación, complejidad de diseño y paciencia (Schoch et al, 2015).
- Esta población se dedicaba a la caza mayor, especialmente de caballos. Muchos huesos correspondieron a grandes mamíferos y exhibían huellas de corte de instrumentos líticos. La acumulación de restos posiblemente corresponde a varios eventos de muerte (Julien et al, 2015; Rivals et al, 2015). Por otra parte, se han recuperado asociados restos de Homotherium latidens (felino de dientes de sable) con evidencias de impactos, lo que sugiere que los Homo utilizaban las lanzas también para defenderse de los carnívoros (Serangeli et al, 2015). Eran hábiles cazadores en la parte superior de la cadena alimentaria, con un alto nivel de planificación y actividades en grupo bien coordinadas y exitosas, probablemente con una división del trabajo (Britt M. Starkovich y Nicholas J. Conard, 2015)
- 1.500 artefactos de piedra en sílex local, utilizadas para el trabajo del cuero y la madera y el corte de carne. Las hachas de mano están ausentes y el conjunto no está relacionado con el Achelense. Presencia de raspadores intensamente retocados, así como piezas denticuladas y con muescas y puntas de escamas gruesas y anchas, con un número más pequeño de formas más delgadas. Las secuencias de reducción son cortas y no se utiliza la tecnología Levallois. (Serangeli y Conard, 2015). Se observa evidencia de enmangamiento en algunas piezas que puede estar relacionada con el hallazgo de mangos de madera en el sitio, pero insuficiente para un diagnóstico inequívoco (Veerle Rots et al, 2015).
- 88 herramientas de hueso, incluidos numerosos fragmentos de huesos largos (en su mayoría de caballo), tres costillas utilizadas como 'retocadores' para afilar herramientas de piedra, y un coxal completo de caballo fue utilizado como yunque para talldo bipolar. La mayoría de los retocadores se prepararon raspando la diáfisis de huesos largos frescos y secos. El reafilado para mantener los filos funcionales, fue exhaustivo. Metapodiales de caballo y bisonte, se interpretan como martillos para alcanzar la médula de otros huesos. Varios martillos metapodiales fueron utilizados además como percutores de talla. Se trata de la evidencia más temprana de herramientas de hueso para usos múltiples (Kolfschoten et al, 2015; Julien et al, 2015).
- Se han recuperado más de 12.000 restos de fauna. La buena conservación del colágeno óseo ha permitido medir las proporciones de carbono y nitrógeno de isótopos estables para diferentes taxones herbívoros. Los resultados sugieren un hábitat abierto (Kuitems et al, 2015)
Vérteszöllös (Hungría). 1965. Occipital y molar deciduo, asociados con fauna e industria clactoniense. 0,475-0,25 Ma con mucha incertidumbre (Cherdyntsev, 1971). El occipital no se parece al de los neandertales y es más primitivo (Thoma, 1966).
 |
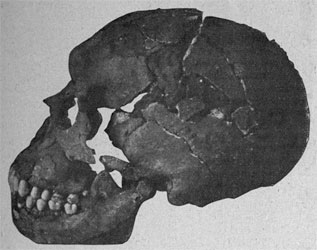
| Cráneo de Ceprano |
CEPRANO Y OTROS EN ITALIA
En Notarchirico (Basilicata, Italia) aparecieron restos líticos Modo 2 datados en 0,6 Ma. En la base hay restos de industrias de Modo 1.
En La Pineta di Isernia fue hallado en 2014 un incisivo deciduo (IS42) en el nivel arqueológico 3 coll (que recubre la capa 3a). Se trata del resto humano más antiguo de Italia, datado por 40Ar/39Ar, entre hace 583-561 ka, a finales de MIS 15 (Peretto et al, 2015). Está asociado a herramientas no achelenses, pese a ser un poco más joven que la difusión del achelense en Europa Occidental, lo que apoyaría la continuidad en refugios de las poblaciones europeas preexistentes.
En La Pineta di Isernia fue hallado en 2014 un incisivo deciduo (IS42) en el nivel arqueológico 3 coll (que recubre la capa 3a). Se trata del resto humano más antiguo de Italia, datado por 40Ar/39Ar, entre hace 583-561 ka, a finales de MIS 15 (Peretto et al, 2015). Está asociado a herramientas no achelenses, pese a ser un poco más joven que la difusión del achelense en Europa Occidental, lo que apoyaría la continuidad en refugios de las poblaciones europeas preexistentes.
 |
| Estratigrafía y hallazgos de Ceprano. Manzi, 2016. |
En 1994 Italo Bidditu halló entre los sedimentos de una cuneta de una carretera en Ceprano, Frosinone, Italia un cráneo que conserva prácticamente toda la bóveda, pero sin base, cuya capacidad podría ser hasta de 1.200 cc. En un principio fue datado en más de 800 ka, pero una revisión de Aurélien Mounier et al, 2011 lo data en 0,430-0,385 Ma. No está asociado a macro o microfauna, restos polínicos ni industria lítica, por lo que hay que pensar en un arrastre.
Debido a la datación dudosa y a sus características peculiares, fue relacionado con muy diferentes posiciones filogenéticas, e incluso le fue asignada una nueva especie: Homo cepranensis (Mallegni et al, 2003), pero actualmente se asigna a heidelbergensis (Manzi, 2016).
Fontana Ranuccio. Dientes datados en 0,458 Ma: dos primeros molares inferiores (izquierdo y derecho), un incisivo inferior izquierdo y un posible canino superior izquierdo. La comparación con otros ejemplares contemporáneos y diacrónicos muestra que los dientes de Fontana Ranuccio son morfológicamente similares a Atapuerca-Sima de los Huesos, Arago XIII y neandertales (Mauro Rubini et al, 2014).
Cava Pompi. Fragmentos de cráneo, cúbito y tibia. 0,45 Ma.
Visogliano. Mandíbula y cinco dientes sueltos. 0,4 Ma.
Vanosa-Notarchirico. Fémur asociado a industria de modo 2. No se ve técnica de Levallois. 0,4-0,35 Ma.
Fontana Ranuccio. Dientes datados en 0,458 Ma: dos primeros molares inferiores (izquierdo y derecho), un incisivo inferior izquierdo y un posible canino superior izquierdo. La comparación con otros ejemplares contemporáneos y diacrónicos muestra que los dientes de Fontana Ranuccio son morfológicamente similares a Atapuerca-Sima de los Huesos, Arago XIII y neandertales (Mauro Rubini et al, 2014).
Cava Pompi. Fragmentos de cráneo, cúbito y tibia. 0,45 Ma.
Visogliano. Mandíbula y cinco dientes sueltos. 0,4 Ma.
Vanosa-Notarchirico. Fémur asociado a industria de modo 2. No se ve técnica de Levallois. 0,4-0,35 Ma.
Castel di Guido. Fragmentos craneales. Una diáfasis de fémur. Asociados a industria modo 2. 0,4 Ma.
Ponte Mammolo (Italia). Un fémur. 0,2 Ma.
Grotta del Principe (Italia). Pelvis de individuo femenino. 0,2 mda.
Grotta del Principe (Italia). Pelvis de individuo femenino. 0,2 mda.
Sedia del Diavolo (Italia). Fragmentos de cráneo y fémur y un metatarsiano. 0,2 mda.
Grotta del Poggio (Italia). Un diente y varios poscraneales. 0,2 mda.
Casal de Pazzi. Fragmento de parietal. 0,2 mda.
Estos hallazgos sugieren la presencia de refugios para la población humana en el centro sur de Italia hasta, al menos, MIS 11 (Peretto et al, 2015).
Estos hallazgos sugieren la presencia de refugios para la población humana en el centro sur de Italia hasta, al menos, MIS 11 (Peretto et al, 2015).
 |
| F. Mallegni (2011) |
BOXGROVE
Boxgrove (East Anglia). Excavado por Mark Roberts en los 90. El yacimiento se mantuvo intacto desde que hace 0,5 Ma fuese cubierto de forma rápida por el fango, lo que permitió su conservación. Se han encontrado huesos de elefante, rinoceronte, caballo, ciervo, león, lobo, hiena, castor, liebre, ratón de campo y rana. Muchos huesos conservan huellas líticas y se ha podido reconstruir el proceso de matanza o despiece. Para Roberts, los animales fueron cazados mientras que Gamble defiende que son fruto de la carroña. Los únicos restos humanos son dos dientes y parte de una tibia, grande y robusta, adaptada al frío, que correspondió a un individuo de unos 1,80 m y 90 kg (0,5 Ma); su morfología es similar a la de los neandertales, pero esto por sí solo no tiene significación filogenética, porque también hay coincidencias con la morfología de otras poblaciones. No hay hogares ni señales de estructuras. Numerosos útiles achelenses, incluyendo bellos bifaces. Los espeleotemas fueron datados por U-series de alta resolución en 600 ka (Bischoff et al, 2007), pero la validez para los restos humanos es discutible (Endicott et al, 2010).ARAGO Y OTROS EN FRANCIA
- Arago 21, Tautavel Man. Descubierto en 1971 por Henry de Lumley en el estrato III. Rostro casi completo, con 5 dientes molares y parte de la bóveda craneal. Su capacidad craneal es de 1.150 cc. Bóveda craneana baja, huesos malares y rostro macizos, pómulos engrosados, maxilar inflado, sin fosa poscanina; parte media de la cara proyectada hacia delante; el frontal es estrecho, la angulación del occipital acusada, parietal grueso y corto; la sección transversal de la bóveda, ligeramente tectiforme; las líneas temporales, altas y marcadas; órbitas relativamente bajas y cuadrangulares, coronadas por fuertes rebordes supraorbitales en doble arco, con depresión glabelar neta.
- Arago 2. Mandíbula de un individuo femenino. Prognatismo.
- Arago 13. Mandíbula de un individuo masculino.
- Dientes de 10 individuos distintos.
- Arago 44. Hueso ilíaco. En conjunto con otros huesos nos permite estimar una talla de 1,64 m.
- Arago 131. Mandíbula con dos dientes, descubierta en 2012.
- Macromamíferos y herramientas líticas.
- En 2013 se hallaron siete dientes, una mandíbula y un fémur.
- En 2015, Arago 149, un incisivo central inferior muy desgastado, en un nivel de ocupación datado en 0,56 Ma.
Vergranne (Francia) canino de menos de 0,5 mda.
Pech-de-l’Aze (Francia). Infantil. 0,4-0,2 Ma.
Orgnac 3 (Francia). Decena de dientes aislados, parecidos a los de Arago. Aparece técnica de Levallois. 0,35-0,32 mda.
 |
| Arago 21 |
AROEIRA
Cráneo dscubierto en la Gruta da Aroeira, Torres Novas, centro de Portugal, datado en hace ca 436-390 ka (Daura et al, 2017). Se trata del cráneo humano más occidental del Pleistoceno Medio europeo y uno de los fósiles humanos más antiguos en un contexto achelense. La presencia de huesos quemados sugiere un uso controlado del fuego.
El espécimen conserva la mayor parte de la mitad derecha de la bóveda craneal (con la excepción del hueso occipital) y el maxilar preserva un fragmento derecho del suelo nasal y dos molares fragmentarios. Su capacidad es de unos 1.200 cc.
El espécimen conserva la mayor parte de la mitad derecha de la bóveda craneal (con la excepción del hueso occipital) y el maxilar preserva un fragmento derecho del suelo nasal y dos molares fragmentarios. Su capacidad es de unos 1.200 cc.
La Gruta da Aroeira deparó entre 1998-2002 un rico conjunto lítico (n = 387) con bifaces achelenses, herramientas retocadas, núcleos, escamas, etc., pricipalmente de cuarcita pero también de sílex. El Método Levallois está ausente. También se recuperaron restos de fauna altamente fragmentados, incluyendo huesos quemados, y dos dientes humanos:
- Aroeira 1. Canino inferior izquierdo. Moderadamente grande, sobre todo en comparación con la muestra de Atapuerca.
- Aroeira 2. Tercer molar superior izquierdo. Uno de los más grandes del Pleistoceno Medio.
En 2013 continuaron los trabajos de campo centrados en la datación de la secuencia, que abarca un espesor de ~ 4 m, y comprende tres grandes unidades. De forma casual se descubrió un cráneo (Aroeira 3) en la base de la unidad 2. Alrededor del cráneo había cementado la brecha y en el hallazgo y la recuperación se dañaron y fragmentaron varias piezas y se produjo un notorio orificio. En conjunto, los dientes y el cráneo corresponden a una edad adulta.
 |
| Aroerira 3, completamente restaurado en vista lateral (A) y reconstrucción virtual y fósil original en vista inferior (B). Barras de escala = 5 cm. |
La combinación de rasgos en el cráneo Aroeira 3 no se ve en ningún otro individuo del Pleistoceno Medio.
- Toro supraorbital continuo y grueso similar al del cráneo Bilzingsleben.
- Apófisis mastoides corta como en la muestra de Steinheim.
- Proceso postglenoideo grande y triangular como en la muestra de Atapuerca (SH).
- Eminencia articular elevada que contrasta con la más plana que generalmente se ve en la muestra de Atapuerca (SH) y en el cráneo de Steinheim. Se ha argumentado que una eminencia articular aplanada es una característica que aparece muy temprano en la evolución neandertal.
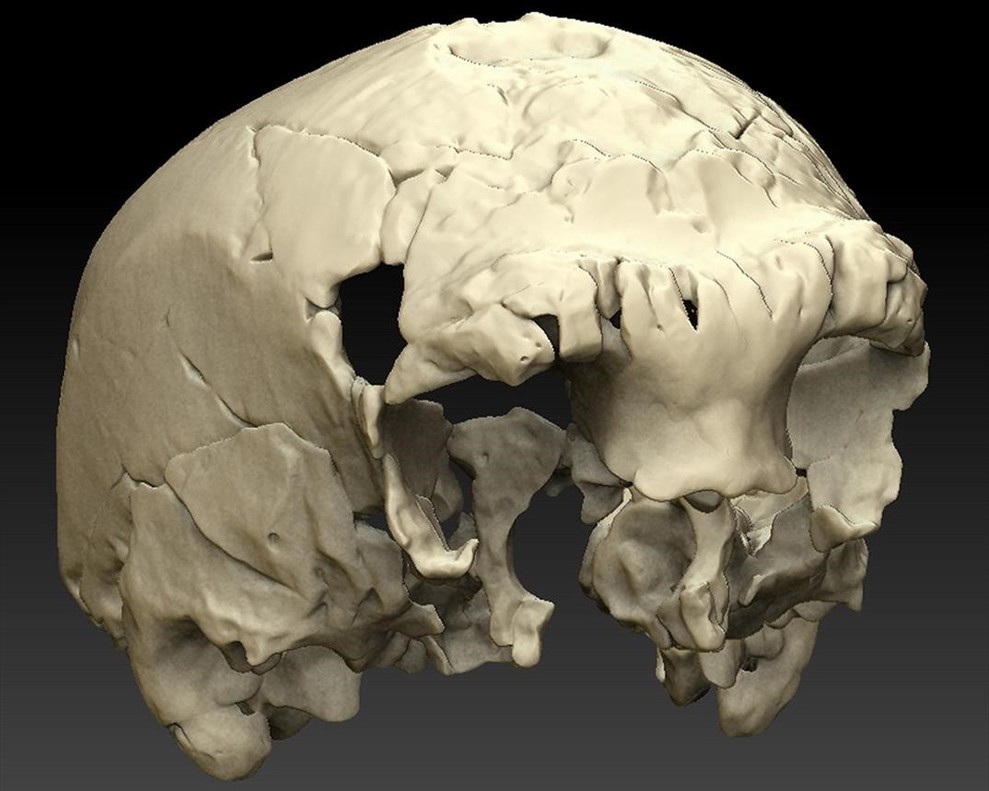 |
| Reconstrucción virtual de Aroeira 3 |
Esta nueva combinación de caracteres apoya, según Zilhão, la idea de una única especie humana en en Europa en este periodo, muy variable y también el Modelo de Fuentes y Sumideros. Según Arsuaga, se trata de un pre-neandertal más primitivo que los homínidos de la Sima de los Huesos.
Bibliografía:
- de Lumley, H., and M. A. de Lumley. "Découverte de restes humains anténéanderthaliens datés du début du Riss à la Caune de l’Arago (Tautavel, Pyrénées Orientales)."Comptes Rendues des seances de l'Acacdemie de Science de Paris 272 (1971): 1729-1742.
- de Lumley, H., and M. A. de Lumley. "PreNeanderthal remains from Arago cave in
southwestern France."Yearb. Phys. Anthropol 17 (1973): 162-168. - Yokoyama, Y., and H. V. Nguyen. "Direct dating by non destructive gamma ray spectrometry of fossil human skull Arago XXI, fossil animal bones and stalagmites of the Caune de l’Arago at Tautavel." Proceedings Pretirage (1981): 355-375.
- Rightmire, G. Philip. "Patterns of hominid evolution and dispersal in the Middle Pleistocene." Quaternary.International 75.1 (2001): 77-84.
- Beck, Curt W. "Archaeometric Clearinghouse."Journal of Field Archaeology 9.2 (1982): 279-280.
- Falguères, Christophe, et al. "New U series dates at the Caune de l'Arago, France." Journal of Archaeological Science 31.7 (2004): 941-952.
- Byrne, Louise. "Lithic tools from Arago cave, Tautavel (Pyrénées Orientales, France): behavioural continuity or raw material determinism during the Middle Pleistocene?."Journal of archaeological science 31.4 (2004): 351-364.
























.png)