 |
| SK 48. Paranthropus robustus. |
Paranthropus robustus, Broom (1938).
2,2–1 Ma. En base al descubrimiento en Kromdraai de TM 1517, el único esqueleto parcial de esta especie conocido hasta ahora (Broom, 1938a, Broom 1938b, Broom 1942 y Broom, 1943). Estos hallazgos fueron también los que motivaron la propuesta del género Paranthropus por Broom en 1938. El escolar Gert Terblanche localizó en 1938 los primeros restos: un cráneo parcial y mandíbula.
- Fragmentos de cráneo incluyendo cinco dientes y pocos fragmentos de esqueleto.
- Único esqueleto parcial de esta especie conocido hasta la fecha.
- Las circunstancias del hallazgo y el contexto estratigráfico son confusos.
- Algunos de los restos que componen TM 1517, se considera actualmente que pertenecieron a cercopitécidos.
La desaparición de la especie pudo estar relacionada con un cambio climático (Caley et al, 2018).
 |
| Ubicación de Kromdraai. Braga et al, 2016. |
Yacimientos:
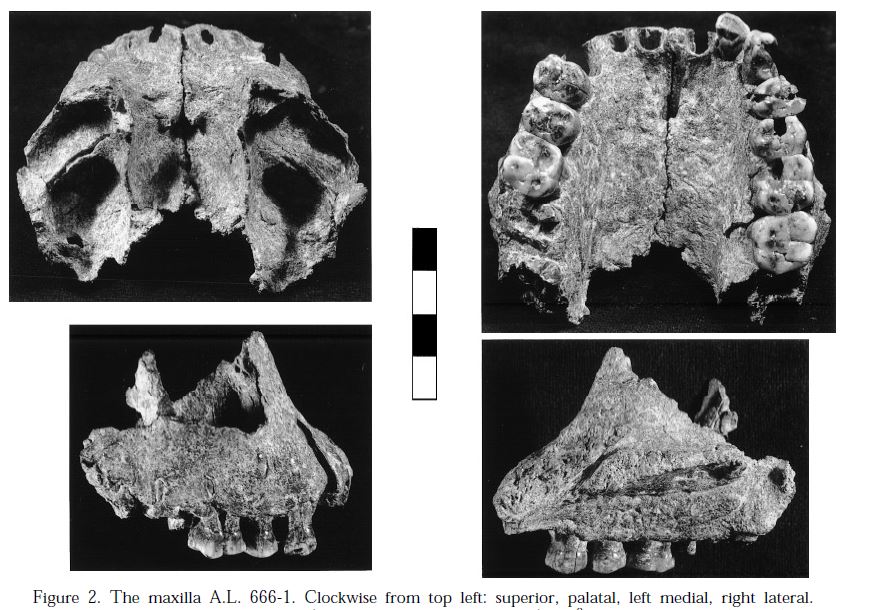
- Kromdraai B (KB) (Gauteng, Sudáfrica). 2-1,5 Ma. Es un vano de dolomita rellenado de depósitos fosilíferos, 2 km al este de Sterkfontein. La datación es imprecisa y, frente a la hipótesis más aceptada de que Paranthropus tiene su origen en Africa del Este con P. aethiopicus, se ha sugerido que al menos algunos especímenes de Kromdraai datan del origen del género Paranthropus (Kaszycka 2002 y Tobias 1988) y son intermedios entre los hominini de Makapansgat Miembros 3/4 y Sterkfontein Miembro 4, por un lado y los más derivados de Swartkrans por otro. Las excavaciones fueron dirigidas por Broom (1938-1944), Brain (1955-1956), Vrba (1977-1980) y Thackeray (1993-2002). Desde 2002, las excavaciones corren a cargo del Kromdraai Research Project. Los contextos para las excavaciones de Broom y Brain son confusos, imposibilitando una datación precisa. Los carnívoros fueron el principal agente de acumulación.
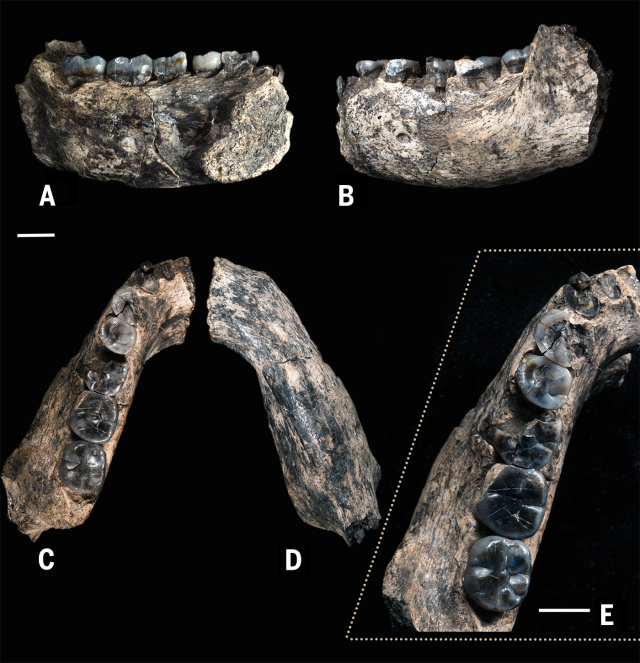
- Individuo 1. Adolescente tardío. TM 1517. Broom (1938a); Broom y Schepers (1946); Broom (1943); Broom (1938b); Day y Thornton (1986); Day (1978); Skinner et al. (2013); Broom (1942).
- Individuo 2. Juvenil. Edad al morir, 1,63-2,02 años (Smith et al, 2015). TM 1536. Fragmento mandibular izquierdo con I1, I2, di2, dm1-2, M1; L dC. Broom (1941); Broom y Schepers (1946).
- Individuo 3. Adulto. TM 1600. Dos fragmentos del corpus mandibular, con M2-M3 (fragmento1) y P3 (fragmento 2). Brain (1981).
- Individuo 4. Juvenil. TM 1601. dm1 inferior derecha, P3, C, P4 germinales inferiores derechos, M1 germinal superior izquierdo, dc inferior derecho. Brain (1981).
- Individuo 5. Adulto. TM 1602. Fragmento maxilar derecho con raíces de P4 a M3. Brain (1981).
- Individuo 6. Adolescente. TM 1603. M3 superior izquierdo. Dump, 1944; Broom and Schepers, 1946.
- Individuo 7. TM 1604. Juvenil. dm2 inferior izquierdo. Brain (1981).
- Individuo 8. Adulto. TM 1605. Cadera izquierda. Brain (1981); Robinson (1972).
- Individuo 9. Adulto. KB 5063. M1 superior derecho. Vrba (1981).
- Individuo 10. Adulto. KB 5163. C inferior derecho. de Ruiter (2004).
- Individuo 11. Juvenil. KB 5222. M3 superior izquierdo. Vrba (1981).
- Individuo 12. Juvenil. KB 5223. dc, dm1-2 inferiores izquierdos; dm2 inferior derecho; incisivos permanentes inferiores; M1 inferior derecho; M1 inferior izquierdo. Vrba (1981), Grine (1982), Braga y Thackeray (2003).
- Individuo 13. Adulto. KB 5226. M3 inferior izquierdo. Vrba (1981).
- Individuo 14. Adulto. KB 5383. M1 superior derecho. Vrba (1981).
- Individuo 15. Adulto. KB 5389. I1 superior izquierdo. de Ruiter (2004).
- Individuo 16. Juvenil. KB 5503. dm2 inferior derecho.Thackeray et al (2001).
- Posiblemente Individuo 1. KB 5522. Fragmento del eje del húmero izquierdo. Thackeray et al (2005).
- Posiblemente Individuo 1. KB 5524. Cara lingual de una corona molar desgastada, posiblemente M1 o M2.Braga et al (2013).
- Individuo 17. Juvenil. KB 6067 es una porción petrosa del hueso temporal de un individuo juvenil. Su morfología lo relaciona con P. robustus, pero también con StW 53 (Homo gautengensis). Sin embargo, el tamaño de la ventana oval es más parecido al de la de algunos especímenes de Sterkfontein Member 4 (Australpithecus africanus). Si KB 6067 se incluye dentro de P. robustus, esto puede representar una condición evolutivamente menos derivada que la deducida a partir de TM 1517 y otros ejemplares conespecíficos. Braga et al (2013).
- Kromdraai E (KE). Excavaciones a partir de 2014 a cargo del Kromdraai Research Project. Hallazgos pendientes de publicación.
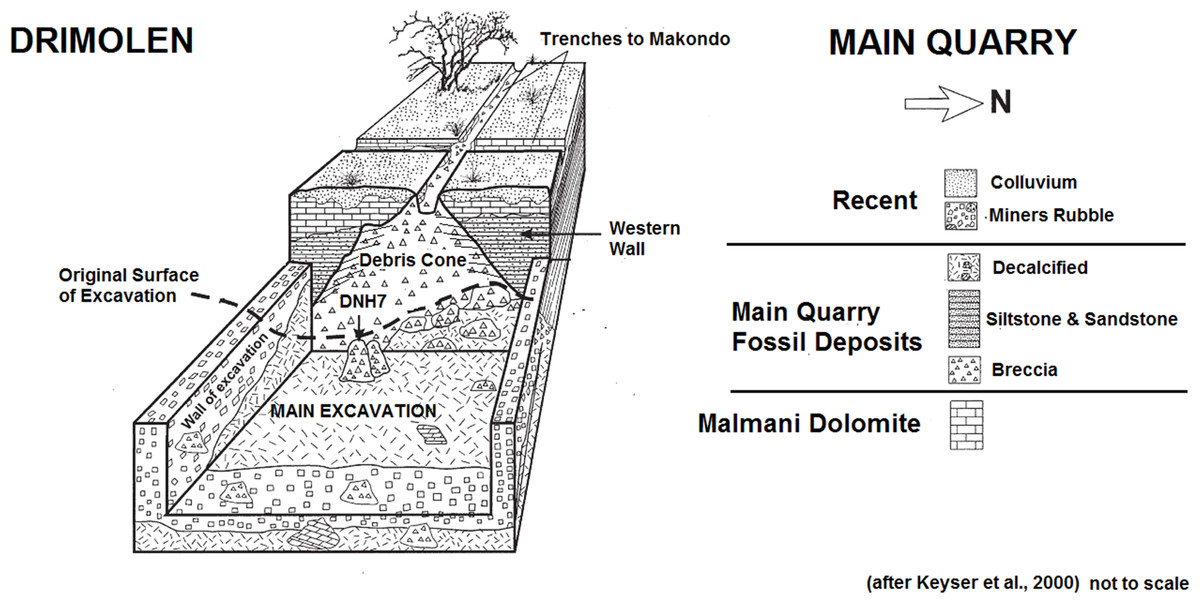 |
| Esquema de las excavaciones de Drimolen. Adams et al, 2016. |
- Sterkfontein Miembro 5. 2,18 Ma (Granger et al, 2015). Algunos dientes.
- Drimolen. 2,04
- -1,5 Ma. El análisis de macromamíferos asociados fue realizado por Adams et al (2016).
- DNH 7, Eurydice. 1994. 1,5-2 Ma. André Keyser. Cráneo casi completo (el más completo conservado de la especie) y mandíbula inferior de una hembra.
- DNH 8 Orfeo. Mandíbula inferior de un macho.
- DNH 44. Mandíbula. Edad al morir, 1,70 años (Smith et al, 2015).
- DNH 47. Maxilar y dientes asociados. Edad al morir, 0.67-0,77 años (Smith et al, 2015).
- DNH 60. Dientes juveniles asociados con un cráneo.
- DNH 84. Maxilar y dientes asociados. Edad al morir, 2,24 años (Smith et al, 2015).
- DNH 101. Molar juvenil aislado.
- DNH 107. Dientes asociados. Edad al morir 4,82 años (Smith et al, 2015).
- DNH 108. Dientes asociados. Edad al morir 5,35-5,53 años (Smith et al, 2015).
- DNH 152. Cráneo de un macho. El más antiguo de la especie en África del Sur (Herries et al, 2020).
 |
| Plano y estratigrafía de Swartkrans. Pickering et al, 2016. |
- Swartkrans. Miembro 1 (2,25-1,7 Ma), Miembro 2 (1,7 Ma) y Miembro 3 (1 Ma) (Pickering et al, 2011; Gibbon et al, 2014). Es el yacimiento con una muestra más amplia de Paranthropus robustus. Los ejemplares de Swartkrans muestran unos dientes mucho más grandes y por ello se ha sugerido que representan una especie diferenciada: Paranthropus crassidens (Broom 1949, Broom 1950, Howell 1978, Grine 1982, Grine 1985 y Grine 1988). Sin embargo, los restos dentales de Drimolen, de tamaño intermedio, favorecen la hipótesis de una única especie (Keyser et al, 2000 y Moggi-Cecchi et al, 2010).
- SK 12. Cráneo. Macho.
- SK 46. Cráneo. Macho.
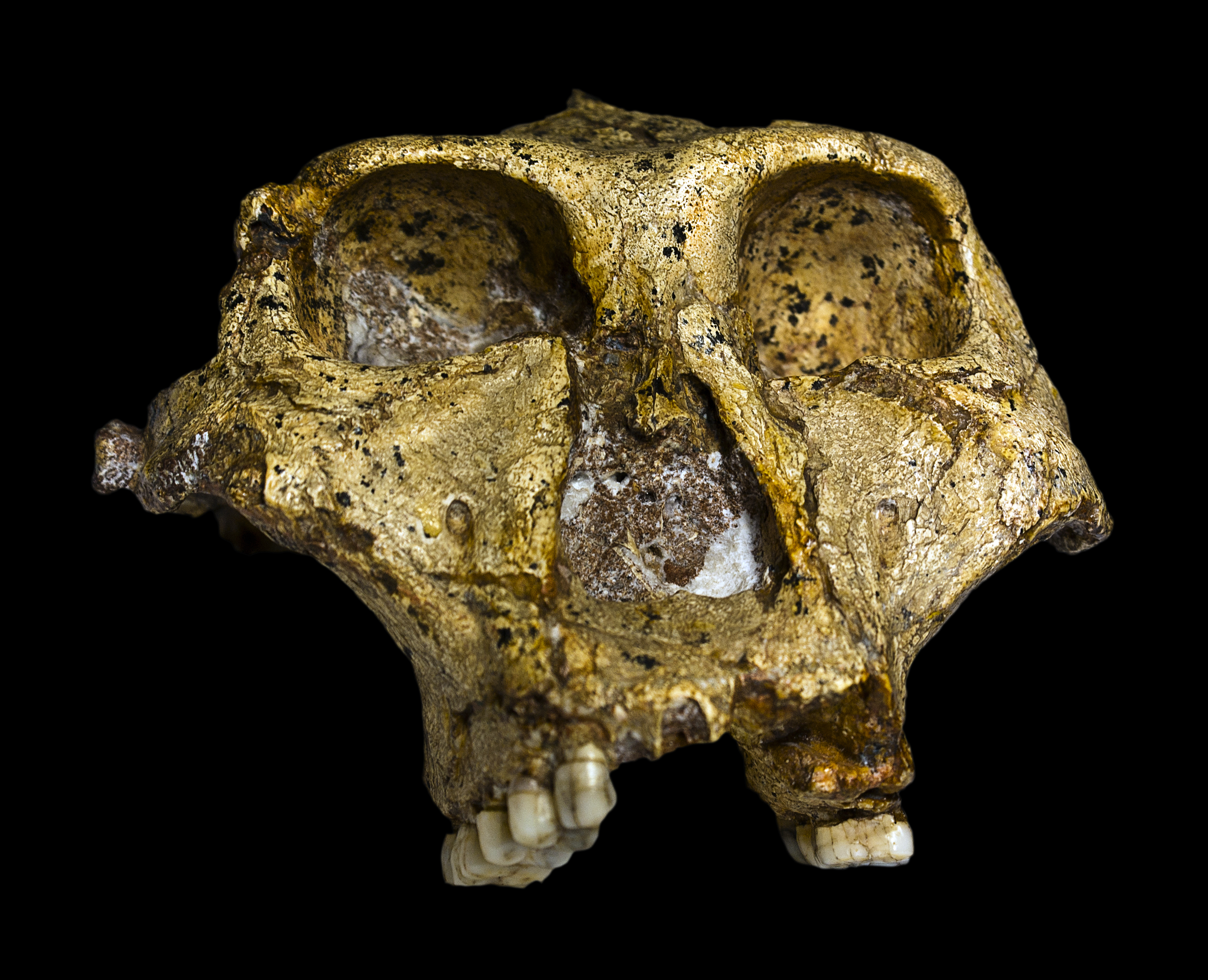
- SK 48. 1950. Mr. Fourie. Se trata de un cráneo, de hembra adulta (macho para Charles A. Lockwood et al (2007). Es el cráneo más completo de los robustus.
- SK 61. Mandíbula.
- SK 62. Mandíbula. Edad al morir, 3,12 años (Smith et al, 2015).
- SK 83 Cráneo. Macho.
- SKX 10924. Lague, 2015. Pequeño fragmento distal de húmero. Asignado por Susman, de Ruiter y Brain (2001) a erectus.
- Varios dientes descritos por Pickering et al, 2016.
 |
| SK 46. Paranathropus robustus |
Estatura 1,32-1,10, un poco superior a la de los australopithecus.
Peso 40-32 kg.
Bipedación casi humana.
Los restos de las manos indican mayor finura táctil que la de los australopitecus.
Según Sandi Copeland (2011), tras estudiar la procedencia de los individuos a partir de rastros en los dientes, los grupos de africanus practicaban la filopatría (los machos se unían a hembras de fuera).
Para Charles A. Lockwood et al (2007)P. robustus muestra un alto dimorfismo sexual y un desarrollo retardado de los machos con respecto a las hembras. Este patrón sugiere que la estrategia reproductora masculino consistía en monopolizar grupos de hembras, de una manera similar a como lo hacen los gorilas de espalda plateada. El costo para los varones debió ser una alta tasa de depredación, lo que justifica la abundancia de machos jóvenes en el registro fósil.
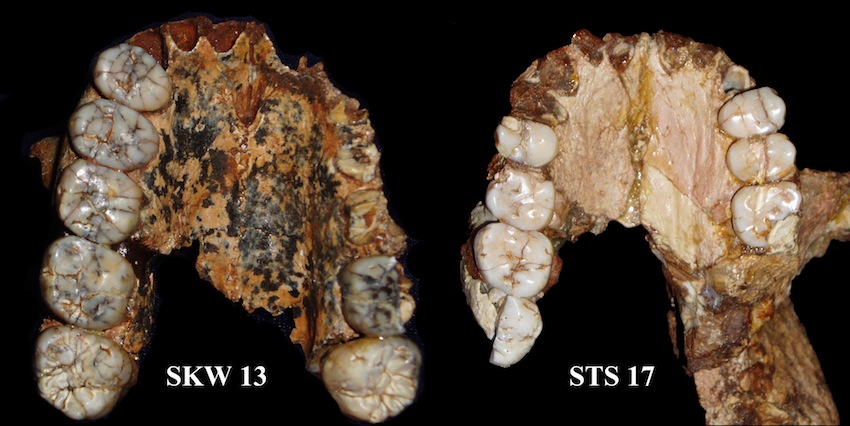 |
| SKW 13 Paranthropus robustus y STS 17 Australopithecus africanus |
Características
- Grandes y anchos molares y premolares. Caninos e incisivos pequeños.
- Mandibulas masivas, cortas, altas y extremadamente robustas con forma de U divergente (mastiación potente). No tanto como boisei. Corpus amplio y profundo y una alta rama, relativamente vertical. No se descarta que, aparte de la dieta vegetal, ingiriese insectos. Según los estudios de Andrew Sillen y Julia Lee-Thorp, P. robustus ingería proteínas animales.
- Caja craneal bastante alta y desarrollada, con cresta sagital.
- Volumen cerebral 530 cc.
- Enorme hueso malar.
- Rostro recto, con arco cigomático muy ancho y robusto. Torus supraorbitario masivo.
- Esmalte dental muy grueso, mucho más que el A. africanus. Según Wynn, Cerling y Sponheimer (2013), su dieta estaba compuesta de alimentos procedentes de árboles y arbustos, pero también de hierbas y juncias, con gran variación entre los individuos. Según Ungar y Hlusko (2016), a pesar de su aparatosa dentadura, solo consumía alimentos duros ocasionalmente.
- Los australopitecinos robustos del Este de África también tienen esmalte muy grueso en los molares.
- Podría ser una sinapomorfia, lo que implicaría una rama robusta común antes de la separación.
- Podría ser un rasgo desarrollado por separado en cada especie, lo que convertiría en dudoso el clado robusto.
- Tamaño y proporciones semejantes a los Australopithecus pero más robustos.
- Oposición del pulgar.
Adam van Arsdale: Paranthropus robusutus
Enlaces:
Evidence of termite foraging by Swartkrans early hominids. Lucinda R. Backwell and Francesco d’Errico (2000)Hominid fossils from Kromdraai: a revised list of specimens discoverd since 1938. Thackeray et al (2001).
Microtomographic archive of fossil hominin specimens from Kromdraai B,South Africa. Matthew M. Skinner, Tracy L. Kivell, Stephany Potze and Jean-Jacques Hublin. 2013
Stretching the time span of hominin evolution at Kromdraai (Gauteng, South Africa): Recent discoveries. Braga et al (2016)
Bibliografía:
- Broom, Robert, and John Talbot Robinson. "Man contemporaneous with the Sawartkrans ape‐man." American journal of physical anthropology 8.2 (1950): 151-156.
- Robinson, John Talbot. "Further remarks on the relationship between “Meganthropus” and australopithecines." American journal of physical anthropology 13.3 (1955): 429-445.
- Sillen, Andrew. "Strontium-calcium ratios (Sr/Ca) of Australopithecus robustus and associated fauna from Swartkrans." Journal of Human Evolution 23.6 (1992): 495-516.
- Lee-Thorp, Julia A., Nikolaas J. van der Merwe, and C. K. Brain. "Diet of Australopithecus robustus at Swartkrans from stable carbon isotopic analysis." Journal of Human Evolution 27.4 (1994): 361-372.
- Keyser, André W. "The Drimolen skull: the most complete australopithecine cranium and mandible to date." South African Journal of Science 96.4 (2000): 189-192.
- Blackwell, Lucinda, and Francesco d'Errico. "Early hominid bone tools from Drimolen, South Africa." Journal of Archaeological Science 35.11 (2008): 2880-2894.
- Moggi-Cecchi, Jacopo, et al. "Early hominin dental remains from the Plio-Pleistocene site of Drimolen, South Africa." Journal of Human Evolution 58.5 (2010): 374-405.
Te puede interesar:
|