 |
| Reconstrucción en base a algunos fósiles de Zhoukoudian |
Se inventarían en China 70 yacimientos.
Se han encontrado en Longgupo (Wushan, Sichuan) fósiles de Hominino (un fragmento de mandíbula con dos dientes y un incisivo) y dos piezas de industria lítica datados en 1,8 Ma (Wanpo, 1995). Wood y Turner (1995) aceptan esta datación mientras que Schwartz y Tattersall (1996) han relacionado los restos con Lufengpithecus. La datación es dudosa.
Yuanmou ha proporcionado dos incisivos datados por paleomagnetismo en 1,8-0,9 Ma.
- Han Fei et al (2015) han datado diecisiete dientes fósiles de mamíferos recogidos de diferentes capas de las unidades C II y C III durante las excavaciones de 2003-2006, mediante ESR y U-series, calculando las fechas mediante un modelo US-ESR.
- Las edades de diez dientes de la pared norte de la unidad C III son consistentes, en general, con una fecha de ca 2,35 Ma.
- Las edades de siete muestras de la pared sur no están de acuerdo con el orden estratigráfico: tres dientes son mucho más jóvenes que los otros cuatro. y proporcionan una edad promedio de ca 2,48 Ma.
 |
| Estratigrafía, dataciones y fauna de Longgupo |
En 1964, un equipo del Instituto de Paleontología de Vertebrados y Paleoantropología de Pekín halló en una pequeña colina de Gongwangling (Lantian, Shaanxi, China) restos de Homo, semejantes a los erectus más antiguos de Java (Wu y Poirier, 1995; Wu et al, 1966). Apareció primero un molar superior aislado y varios meses más tarde grandes fragmentos de la bóveda craneal. También se recuperó una gran cantidad de material de fauna de mamíferos. Un predominio de especies tropicales y subtropicales ha sugerido un clima más cálido que hoy (Gu y Jablonski, 1989; Qi, 1989). Se han hallado artefactos líticos (Dai, 1966; Tai y Hsu, 1973). Los materiales óseos de Homo fueron descritos por Woo (1965) y Wu y Poirier, 1995. Los fósiles humanos (PA 1051-6) incluyen un frontal completo, gran parte de los parietales, la mayor parte del temporal derecho, parte de los nasales, una gran parte del maxilar derecho con segundos y terceros molares asociados, y parte del maxilar izquierdo. La preservación de los fragmentos de hueso es extremadamente pobre. Superestructuras muy marcadas: torus frontal y occipital, gran espesor de los gruesos del cráneo, constricción postorbitaria marcada, boveda craneal alargada y baja. 770 cc.
Zhao-Yu Zhu et al (2014) han datado los fósiles en ca 1,62 Ma, probablemente ca. 1,63 Ma, lo que hace de Gongwangling el segundo sitio más antiguo fuera de África con restos craneales (después de Dmanisi).
En Yunxian (Hubei) se realizaron los siguientes hallazgos, datados en 1,1-0,9 Ma (Vialet et al, 2011). Ambos cráneos están muy deformados, son muy robustos y tienen un perfil bajo y alargado. Dennis A. Etler ha destacado las similitudes con ATD6-69 (Atapuerca, Gran Dolina):
· 1989. Yunxian I. Cráneo.
· 1990. Yunxian II. Craneo de 1.100 cc asociado a restos de fauna e industria lítica.
En Chenjiawo (Lantian) se halló Lantian 1, una mandíbula carente de las últimas muelas, datada por estratigrafía en 0,65 Ma.
En la cueva Peking Man Sitede Zhoukoudian (Pequín), excavada por J.G. Andersson, entre 1929-1937 se encontraron 5 bóvedas craneales, 14 mandíbulas fragmentarias, 147 dientes aislados y huesos de esqueletos, que fueron inicialmente atribuidos al Sinanthropus pekinensis. La cueva fue ocupada por Homo y hienas de forma intermitente.
Los restos podrían tener entre 0,55 y 0,3 Ma. En todo este tiempo, se constata una gran estabilidad evolutiva, indicio del aislamiento de esta población.
Los restos más completos corresponden a:
- Skull III, descubierto en Locus E en 1929 . Es el cráneo de un adolescente con capacidad craneana de 915 cc.
- Skull II, descubierto en Locus D en 1929, sólo reconocido en 1930, es un adulto o adolescente con capacidad de 1030 cc.
- Skulls X, XI and XII (antes llamados LI, LII y LIII) fueron descubiertos en Locus L en 1936. Se trata de un adulto hombre, una hembra y un jóven, con capacidades de 1225 cc, 1015 cc y 1030 cc respectivamente. (Weidenreich 1937).
- Skull V: dos cráneos descubiertos en 1966 y otros fragmentos descubiertos en 1934 y 1936 de bóvedas craneanas con una capacidad de 1140 cc. (Jia and Huang 1990).
- En 2015 se comunicó la identificación de un canino de adulto hembra desgastado y rasgado procedente de este yacimiento en una caja en el Museo de Uppsala.
La industria lítica asociada fue estudiada por Teilhard de Chardin y W. C. Pei. Carece de los utensilios de gran formato que definen el Modo 2, pero en cambio contine otros útiles próximos al Modo 3.
Se hallaron restos de hogares y huesos quemados, quizá como combustible, pero no está claro su origen.
Llama la atención la ausencia de huesos poscraneales lo que sugiere algún tratamiento simbólico. En algún cráneo hay marcas de corte que sugieren canibalismo.
Los cráneos son alargados, bajos, con la máxima anchura a nivel de los huesos temporales, occipital muy angulado, paredes óseas gruesas; frontal bajo y huidizo. Capacidades craneles entre 915-1.225 cc. Se ha sugerido la necesidad de agrupar los restos de Zhoukoudian con los de Java, en virtud de sus semejanzas morfológicas (Weindenreich 1938, agrupacionistas). Las diferencias en la dentición y en detalles del cráneo se interpreta por ellos como una evolución lógica justificada por la mayor antigüedad de los restos de Java. Para Emiliano Auirre (2008), el punto punto de vista agrupacionista impide valorar sus rasgos verdaderamente progresivos: un cráneo más alto y ancho, frente más elevada y rostro retraído.
| Segundo y tercer molar inferior (PA834-1 y PA834-2) de la Cueva Longtan. (o: oclusal, b: bucal, m: mesial, l: lingual, d: distal). Planos de sección sagital de PA834-1 (V) y PA834-2 (VII). Sección transversal de PA834-1 (VI) y PA834-2 (VIII) en el nivel indicado por las líneas rojas. I-VIII no se escalan. |
Para Howell estos restos constituyen el paleodemo ZKD, junto a la mandíbula de Chenjiwao y otros hallazgos, a los que se han ido incorporando los siguientes:
- Halu, Tangshan (Nanjing). 1993. Nanjing 1 y Nanjing 2. Dos cráneos fragmentarios entre 0,62-0,58 Ma.
- Yiyuan. Fragmento craneal y dientes. 0,4-0,3 Ma.
- El estudio comparativo métrico y morfológico realizado por Song Xing et al (2016) apoya una atribución humana y relaciona los dientes de Yiyuan con las muestras de Zhoukoudian, Chaoxian y Hexian. Estas muestras son menos derivadas que las de Panxian Dadong (sapiens arcaico), y son también diferentes de las del Pleistoceno Temprano de Indonesia, lo que apoya la idea de que la taxonomía de los homínidos del Pleistoceno de Asia se ha simplificado en exceso.
- Xinghuasan, Longushan, Builongdong, Loggudong y Huludong. Dientes. 0,4-0,3 Ma.
- Chaoxian. Un occipital y un maxilar con seis dientes datados en 0,2-0,16 Ma.
 |
| Hexian I. Cui y Wu, 2015. |
- Cueva Longtan (Longtandong), Hexian, Anhui, Este de China: 1980-1982. Calota casi completa, dos fragmentos craneales, corpus mandibular izquierdo parcial con M2 y M3, diez dientes aislados y restos poscraneales fragmentarios de cuatro individuos, datados en 0,412 ± 25 ka (Huang et al, 1981; Grün et al, 1998). Fauna asociada pero no artefactos.
- Hexian I: neurocráneo hallado en 1982 (Cui y Wu, 2015).
- Los dientes son primitivos métrica y morfológicamente y se superponen con los de H. ergaster y Hominini tempranos del Este de África en sus grandes dimensiones y complejidades oclusales (Song Xing et al, 2014).
- Xichuan. 13 dientes aislados recuperados en tiendas de medicina tradicional. Sin datación.
- Tangshan. Dos cráneos fragmentarios datados por U-series en ca 0,5-0,4 Ma.
- Penghu 1 es un lateral derecho mandibular con los premolares y molares intactos, pero muy desgastados, en su lugar. Se conservan los alvéolos de los incisivos y caninos. El proceso coronoides está roto. No hay signos de deformación postmortem o abrasión de la superficie ni de, ni signos de patologías; M3 es congénitamente ausente. Fue dragado por una red de pesca en el canal submarino Penghu a una profundidad de -60 a -120 m, a 25 km de la costa occidental de Taiwán, junto con otros fósiles de vertebrados conocidos como fauna Penghu. El canal había sido parte del continente asiático durante el Pleistoceno. El nivel del mar disminuyó más de 60 m entre hace 190-130 ka y hace 70-10 ka. Penghu 1 probablemente se depositó por la acción fluvial durante una de estas fases. Existieron eventos más antiguos de bajo nivel del mar, pero la fauna asociada no se corresponde con esa antigüedad. Por lo tanto, Penghu 1 es más joven que 450 ka, y muy probablemente entre 190-130 ka o 70-10 ka. Estas fechas son inesperadamente tardías para la apariencia robusta y primitiva de Penghu 1, desconocida en el registro asiático con excepción de Hexian (Chun-Hsiang Chang et al, 2015).
 |
| Dientes de Hexian. María Martinón-Torres et al (2016). |
En base al estudio de los dientes de Hexian, Song Xing et al (2014) sugieren escenarios evolutivos complejos de convivencia y / o supervivencia de diferentes linajes en Eurasia. Hexian podría representar la persistencia en el tiempo de un H. erectus que habría conservado rasgos primitivos que se perdieron en otras poblaciones asiáticas como Zhoukoudian o Panxian Dadong. Emplazan a futuros estudios explorar si la variabilidad morfológica de las poblaciones de Asia puede ser explicada por un modelo de "fuente-sumidero". Las regiones semiáridas probablemente contenían las poblaciones fuente que se expandirían a regiones áridas colindantes cuando los niveles de población o / y las condiciones climáticas fuesen propicias. La inestabilidad climática y el cambio ambiental en Asia podrían haber favorecido un patrón de fragmentación y aislamiento con alta variabilidad morfológica del registro de Hominini de Asia y la posibilidad de la persistencia de los linajes de Hominini primitivos a través del tiempo.
- Los dientes de Hexian son primitivos métrica y morfológicamente y se superponen con los de H. ergaster y Hominini tempranos del Este de África en sus grandes dimensiones y complejidades oclusales. Zhoukoudian es morfológicamente más derivado, pese a ser contemporáneo o ligeramente más antiguo (Shen et al., 2009). Hexian podría ser también más primitivo que Chaoxian y manifestaría la persistencia en el tiempo de un grupo de erectus que podría haber retenido características primitivas perdidas en otras poblaciones contemporáneas.
Chun-Hsiang Chang et al (2015) proponen varios modelos para explicar la persistencia de Penghu 1, con una morfología tan arcaica, hace menos de 190 ka.
- La retención de características de los Homo tempranos de Asia.
- Debido a que los erectus de Java y China muestran corpus más delgado corpus y molares más pequeños, esta hipótesis implica la presencia de otro linaje en Asia que se prolongó desde el Pleistoceno Temprano.
- Una migración de un grupo africano con mandíbula robusta, posiblemente trayendo consigo la tecnología lítica achelense, alrededor del final del Pleistoceno Inferior, que más tarde evolucionó a nivel local.
- Esto es compatible, pero diferente de una sugerencia reciente de que el Homo arcaico tardío chino recibió alguna influencia genética del oeste y puede ser incluido en H. heidelbelgensis. Penghu 1 parece ser más primitivo y similar a Hexian cuya morfología craneal es distinta de Jinniushan y otros Homo arcaicos tardíos chinos.
- Ambas hipótesis ponen en duda la idea tradicional de que H. erectus fue la única especie de Hominini en el continente asiático en el Pleistoceno Inferior y principios del Pleistoceno Medio, que exhibió variación geográfica clinal en todo el continente desde Indonesia hasta el norte de China.
- Una evolución a partir de una población de erectus de mandibula grácil.
- Esto es contrario a la tendencia general de reducción del tamaño dental en el Homo del Pleistoceno.
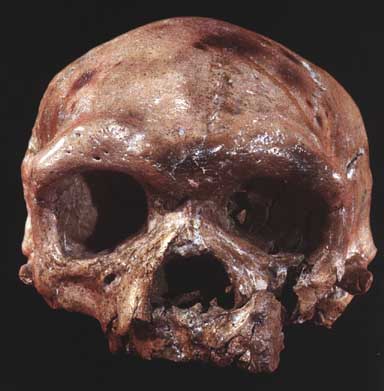 |
| Cráneo de Dali |
Otros hallazgos fósiles en China han descubierto fósiles más modernos que apoyan la coexistencia en esa zona de Homo erectus y Homo sapiens, o al menos la evolución asiática de erectusa sapiens. Para Howell constituyen un paleodemo diferenciado. Restos que se asimilan por su parecido y capacidad craneal al sapiens se han datado en unos 0,58 Ma, lo que supondría que el sapiens asiático ha evolucionado antes que los sapiensEuropeos. Estos hallazgos apoyarían la teoría de la evolución multirregional del sapiens en contra de la teoría Out of África). Sin embargo quedan muchas dudas de que estos fósiles asiáticos puedan ser considerados verdaderos sapiens.
- Dali. 1978. Cráneo completo datado en 0,3-0,25 Ma por U-sereis. Parietal más alzado, con fosas poscanina y maxilar pronunciadas, concavidad en el pómul y rostro corto y vertical, muy poco prognato. Con rasgos parecidos a los restos hallados en Zhoukoudian pero también con rasgos modernos. 1.150 cc.
- Miahoushan. Restos datados entre 0,22 y 0,14 Ma.
- Changyang
- Localidad 4 de Zhoukoudian
- Yanghuidong (Tongzi, Guizhou). Dientes y fragmentos craneales. 0,2 Ma.
- Hathnora (Narmada, India). Cráneo datado en 0,2 Ma. Presenta un mosaico de caracteres de erectus y sapiens. Bóveda craneal más elevada y superestructuras menos marcadas.
- Dingcun. 0,11 Ma.
- Hualongdong. (Jiandé, Dongzhi, Anhui). 0,09 Ma.
- El 11 de octubre de 2015 apareció un cráneo (Dongzhi Man), junto con una variedad de instrumentos de piedra, dientes y fragmentos de hueso humano de al menos cuatro individuos, uno de ellos infantil, así como más de 6.000 fósiles óseos de vertebrados. Se han datado entre 0,412-0,150 Ma.
- Guojiujan.
- En la Región de las Tres Gargantas se han localizado 10 yacimientos de Hominini y otros 30 líticos (Modo 1) y con otras evidencias de actividades humanas. Yacimientos más importantes:
- Yandunbao.
- Miaogou.
- Chibaling.
- Ranjialukou.
- Gaojiazhen.
- Fanjiahe.
- Wuyang.
- Cueva Xinglong.
- Cueva Sunjia.
- Cueva Zhongjiawan.
- Jingshuiwan.
- Zaoziping.
- Dadiping.
- Outang.
- Cueva Leiping.
- Cueva Migong.
- Cueva Huanglong. 2004-2006. 7 dientes asociados a herramientas de piedra y hueso y a fósiles de mamíferos. Datados en 0,1 Ma.
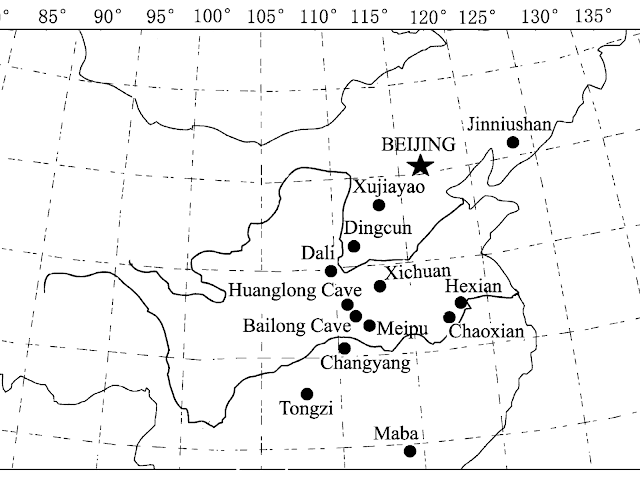 |
| Principales yacimientos de China |
Howell distingue un paleodemo adicional para:
- Jinniusham (Liao Ning, Manchuria). Esqueleto femenino incompleto con el que Howell definió el paleodemo, único hallado en Asia correspondiente al Pleistoceno Medio. 1984. Datado por ESR en 0,28 Ma. Gran anchura de la pelvis. Pudo pertenecer a un individuo de 168 cm y 78 kg. El cráneo, de 1.330 cc, conserva rasgos primitivos, como la bóveda craneana todavía baja y alargada hacia la parte posterior; nariz chata. Rosenberg piensa que la robustez y anchura son adaptaciones a climas fríos. Sin embargo, estos rasgos también están presentes en una pelvis de la Sima de los Huesos de Atapuerca de la misma datación. Por ello Arsuaga et al (1999) creen que estamos ante la morfología primitiva y que solo los sapiens presentan una pelvis estrecha y una forma grácil.
Fuentes y Sumideros. Un modelo para el poblamiento de Asia Oriental.
El conjunto de la Sima de los Huesos presenta una morfología derivada neandertal más pronunciada que los ejemplares de Mauer, Arago o Ceprano. Para explicar esta variabilidad, Dennell, Martinón-Torres y Bermúdez de Castro (2011), Bermúdez de Castro y Martinón-Torres (2013), propusieron un modelo de población que se basa en fuentes y sumideros demográficos. Un pequeño número de fuentes en el sur del continente repueblan áreas más al norte durante los interglaciares, dando lugar a grupos sumidero. Estos grupos sumidero se extinguirían durante las épocas glaciares. De esta forma la variabilidad vendría favorecida por sucesivas dispersiones, fragmentaciones y recombinaciones de las poblaciones.
Algunos estudios han apuntado que el el aislamiento geográfico es uno de los mecanismos que podrían explicar la descontinuidad, la evolución regional y las diferencias morfológicas entre poblaciones dentro de China y entre las de Asia continental y Java (Kaifu, 2006; Zanolli, 2013).
María Martinón-Torres et al (2016), a partir de las evidencias dentales recuperadas en el Este de Asia, consideran que el Modelo de Fuentes y Sumideros explica también el asentamiento humano en Asia Oriental durante el Pleistoceno.
- La variación morfo-dimensional de los ejemplares, no puede ser alojada dentro de un mismo linaje ni a lo largo de una secuencia cronológica y apoya un patrón de ocupación intensa pero discontinua. La fragmentación y el aislamiento habrían favorecido la retención de características primitivas en ciertos grupos y el desarrollo de caracteres más derivados en otros, con diferentes tendencias evolutivas de forma simultánea. Un linaje primitivo pudo coexistir en el tiempo con sapiens (Chang et al, 2015; Curnoe et al, 2012) y cabe especular sobre su posible asimilación y mezcla (Hipótesis Multirregional). Los últimos estudios sobre ADN antiguo, soportan escenarios con la persistencia de linajes antiguos e introgresiones.
- Las oscilaciones climáticas severas, la extensión de las zonas desérticas y la posibilidad de intercambios intermitentes con el las regiones insulares del Este de Asia, en función de cambios en el nivel del mar, son factores clave para identificar los posibles fuentes y sumideros en la región.
- Estudios recientes apuntan a los desiertos como elementos clave que impidieron el asentamiento y la dispersión libre de los Homo por el continente asiático (Dennell, 2013a, 2013b). Es lógico suponer que las expansiones y contracciones de los desiertos a lo largo del Pleistoceno (Liu et al, 1999) tuvieron un impacto importante. Además, los Homo eran particularmente sensibles a la variación en las lluvias (Dennell, 2009), por lo que los monzones probablemente jugaron también un papel importante.
- El análisis del patrón de asentamiento en la cuenca del Nihewan y la adyacente meseta de Loess, sugiere una ocupación intermitente, en los períodos cálidos favorables (Dennell, 2013b).
- Las regiones semiáridas probablemente contenían las poblaciones fuente que se expandirían a regiones áridas colindantes cuando los niveles de población o / y las condiciones climáticas fuesen propicias para la reducción de las áreas desérticas. La inestabilidad climática y el cambio ambiental en Asia podrían haber favorecido un patrón de fragmentación y aislamiento con alta variabilidad morfológica del registro de Homo de Asia y la posibilidad de la persistencia de los linajes primitivos de Homo a lo largo del tiempo.
- Otra cuestión importante es el potencial de las islas para actuar como fuentes o sumideros dependiendo de los cambios en el nivel del mar y la existencia de puentes de tierra con Asica continental (Dennell et al, 2014; Louys y Turner, 2012). La presencia de Hominini en Flores o Taiwan constituyen ejemplos del papel demográfico que pueden representar las islas.
Bibliografía:
- Weidenreich, Franz. "The skull of Sinanthropus pekinensis: a comparative study on a primitive hominid skull." (1943).
- Wang, Y. Y., et al. "Discovery of the fossil Dali Man in Shaanxi Province and the preliminary study." Chinese Science Bulletin 24 (1979): 303-306.
- Wu, X. Z. "A well-preserved cranium of an archaic type of early Homo sapiens from Dali, China." Scientia Sinica 24.4 (1981): 530.
- Pope, Geoffrey G. "Craniofacial evidence for the origin of modern humans in China." American Journal of Physical Anthropology 35.S15 (1992): 243-298.
- Etler, Dennis A. "The fossil evidence for human evolution in Asia." Annual Review of Anthropology (1996): 275-301.
- Weiner, Steve, et al. "Evidence for the use of fire at Zhoukoudian, China."Science 281.5374 (1998): 251-253.
- Goldberg, Paul, et al. "Site formation processes at Zhoukoudian, China."Journal of Human Evolution 41.5 (2001): 483-530.
- Shen, Guanjun, et al. "Age of Zhoukoudian Homo erectus determined with 26Al/10Be burial dating." Nature 458.7235 (2009): 198-200.
- Wu, Xinzhi, and Sheela Athreya. "A description of the geological context, discrete traits, and linear morphometrics of the Middle Pleistocene hominin from Dali, Shaanxi Province, China." American journal of physical anthropology 150.1 (2013): 141-157.
Te puede interesar:
|